
Varanus bangonorum, Welton, Luke J., Travers, Scott L., Siler, Cameron D. & Brown, Rafe M., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3881.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:62DB7048-70F2-4CB5-8C98-D7BCE48F4FC2 |
|
DOI |
https://doi.org/10.5281/zenodo.6139291 |
|
persistent identifier |
https://treatment.plazi.org/id/6D728781-FFC1-3A10-C4C7-FF3DFB99A65D |
|
treatment provided by |
Plazi |
|
scientific name |
Varanus bangonorum |
| status |
sp. nov. |
Varanus bangonorum sp. nov.
Figs. 3 View FIGURE 3 , 6–9 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9
Holotype. PNM 9798 (formerly University of Kansas Natural Biodiversity Institute [KU] 335744; Field no. RMB 17757), sub-adult male, collected by SLT and RMB, 16 July 2013, 230 m above sea level (12.787806, 120.915972; WGS-84), Sitio Aruyan, Barangay Malisbong, Municipality of Sablayan, Occidental Mindoro Province, Mindoro Island, Philippines.
Paratopotypes. KU 335742 (RMB Field No. 17720), juvenile, collected 11 July 2013.
Paratypes. KU 335743 (RMB Field No. 17824), juvenile, Lake Libuao, Barangay Malisbong, Municipality of Sablayan, Occidental Mindoro Province, Mindoro Island, Philippines; PNM 9799 (formerly KU 308403), 308437 (ELR Field Nos. 843, 877), juvenile and adult female, Sitio Ulasan, Barangay Harrison, Municipality of Paluan, Occidental Mindoro Province, Mindoro Island, Philippines; KU 305151, 305161, 305163 ( CDS Field Nos. 714, 715, 648), juveniles, Barangay Tinogboc, Municipality of Caluya, Antique Province, Semirara Island, Philippines.
Other Material. See Specimens Examined Section
Etymology. The specific epithet, bangonorum , is a plural noun derived from the name of the indigenous Bangon peoples of Mindoro Island, who reside along the rivers and central Cordillera mountain system in Oriental Mindoro Province. Anthropologists have shown the Bangon tribal group to be culturally distinct from other tribal groups inhabiting Mindoro ( Tweddell 1970; Mangyan Heritage Center). Suggested common name: Bangon Monitor Lizard.
Diagnosis. Varanus bangonorum can be distinguished from congeners by (1) distinct, relatively large dark spots or blotches in the gular region; (2) low counts of dorsal gular scales; (3) low counts of dorsal trunk scales; and (4) low counts of scales around the neck anterior to the gular fold. Additionally, this distinct lineage is biogeographically circumscribed on the deepwater islands of Mindoro and Semirara.
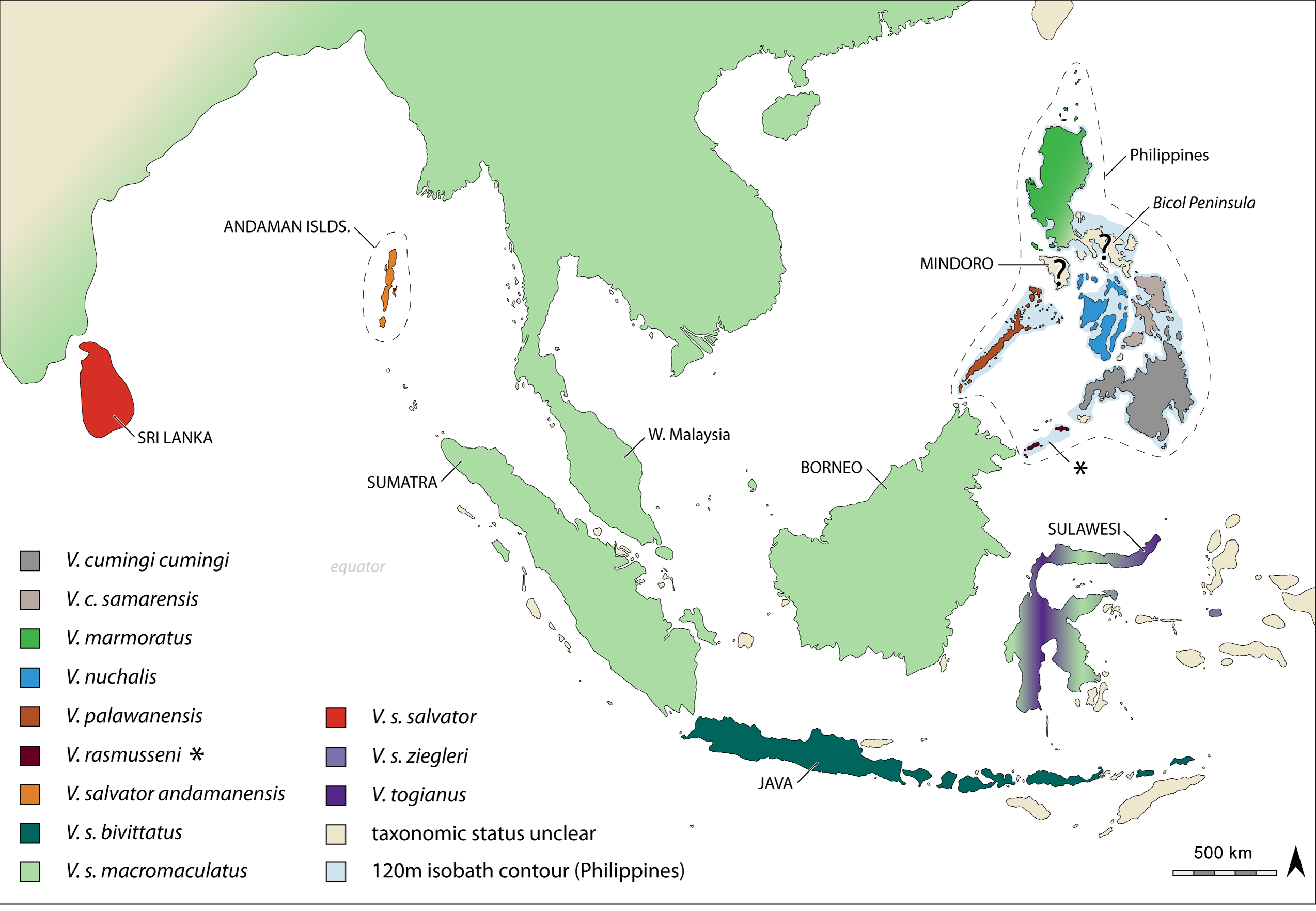
Comparisons. This new species in phenotypically most similar to Varanus marmoratus , from which it can be distinguished by the presence of distinct, dark and irregularly-shaped spots or blotches in the gular region (versus speckling and transverse bands; Fig. 3 View FIGURE 3 ), and an allopatric distribution on the islands of Mindoro and Semirara (versus Luzon and Lubang islands, and the Batanes and Babuyan island groups; Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ). Additionally, although the new species is phenotypically similar to V. marmoratus , it is not closely related to this species ( Fig. 2 View FIGURE 2 , phylogeny). Varanus bangonorum can be distinguished from its closest but allopatric relative, V. palawanensis , by having generally fewer rictus–rictus scales (mean = 56 ± 4, versus 62 ± 4), fewer dorsal scales from the tympanum to the gular fold (mean = 26 ± 2, versus 32 ± 3), fewer dorsal scales from the nuchal fold to the hind limb insertion (mean = 80 ± 9, versus 96 ± 3), fewer total dorsal scales (mean = 106 ± 10, versus 128 ± 3), fewer scales around the neck anterior to the gular fold (mean = 76 ± 6, versus 87 ± 4), and by having distinct, dark irregular spots or blotches in the gular region (versus irregular speckling with anterior transverse bands). Of the remaining geographically proximate species of the V. salvator Complex, this new species can be distinguished from V. nuchalis by a distribution on Mindoro and Semirara islands (versus Visayan islands), generally fewer midbody scales (mean = 136 ± 9, versus 150 ± 7), generally more dorsal scales from the tympanum to the gular fold (mean = 26 ± 2, versus 23 ± 2), and the presence of distinct dark spots or blotches in the gular region (versus a uniform dark gular coloration). Additionally, this new species can be distinguished from the allopatric Bicol species V. dalubhasa by having distinct, irregularly shaped spots or blotches (versus variable speckling and faint anterior transverse bands) in the gular region. Lastly, V. bangonorum exhibits entirely unique haplotypes or haplotype networks, relative to all other members of the V. salvator Complex (see Welton et al. 2010a, b). We have constrained our morphological analyses to the geographically most proximate taxa due to previous studies (Koch et al. 2007, 2010b) demonstrating their distinctiveness relative to the remaining diversity within the V. salvator Complex.
Description of holotype. A sub-adult male, hemipenes not everted; snout–vent length 343 mm; tail 601 mm; head relatively slender, length 65.9 mm, width 38.2 mm, and depth at eye 28.5 mm; snout length 34.0 mm, 51.6% head length; snout rounded anteriorly; narial openings 4.3 mm, ovular, posteriorly elevated, encircled by 9L/9R small polygonal scales; snout elevated sharply just anterior to narial region, then continuing gradually to ocular region; narial region only moderately elevated above horizon of snout; cranial table squarish, slightly wider than long.
Head scales ovular to polygonal ( Fig. 9 View FIGURE 9 ), heterogenous in size with the smallest occuring on lateral margins of cranial table and directly surrounding pinneal scale, and largest occuring dorso-medially from tip of snout to pineal region; supraoculars 7L/6R, subrectangular and elongated transversely; supralabials 52, slightly larger than head scales; infralabials 52, smaller than supralabials; both supra- and infralabials increasing in size anteriorly; nuchals large, circular, generally homogenous in size throughout nuchal region except for directly posterior to cranial table; lateral nuchals small and granular, approximately ¼ the size of dorsals; sutures between scales larger than those on head; nuchals 25, in semi-regular transverse rows from posterior margin of tympanum to forelimb insertion; dorsal trunk scales arranged in 77 semi-regular transverse rows from fore- to hindlimb insertions, ovular, generally smaller than nuchals, and decreasing in size posteriorly to hindlimb insertion as well as laterally, with smallest scales occurring adjacent to limb insertions; midbody scales 131; axilla-groin distance 157 mm; dorsal limb scales ovular, decreasing in size distally and ventrally; fore- and hindlimbs 55 and 82 mm, respectively, 16.0% and 23.9% snout–vent length, respectively, 24.2 and 36.8 mm diameter at insertions, respectively; limb scales present in semiregular rows; scales of manus and pes smaller and more irregular in shape, ranging from circular to nearly square dorsally; supradigitals rectangular dorsally, ovular laterally, with terminal scale nearly twice as large as others; digits terminate in long, slender recurved claws; caudals ovular, small and granular anteriorly, becoming rectangular posteriorly; dorsal keel composed of paired, posteriorly elevated scales commenses 37.7 mm from hindlimb insertion, continuing to tail terminus with scales gradually decreasing in size; caudal scales moderately keeled, in regular transverse and longitudinal rows laterally, decreasing in size posteriorly.
Ventrals less variable than dorsals; gular scales generally rectangular anteriorly, bcoming more ovular towards gular fold; smallest scales medially in anterior third of gular region, with largest scales just anterior to gular fold but followed posteriorly by three transverse rows of smaller, more granular gulars; scales around the neck anterior to the gular fold 67, mid-gular 76; gular scales in 70 irregular rows from tip of snout to gular fold; ventrals of fore- and hindlimbs circular to polygonal, with those of forelimbs slightly smaller than those of hindlimbs; precloacals similar to those of hindlimbs; ventral trunk scales from gular fold to hind limb insertion 83, ovular anteriorly, becoming rectangular posteriorly, present in semi-regular rows; scales decreasing in size laterally and at hindlimb insertions; ventral caudal scales rectangular, smallest just posterior to cloacal opening; scale surface slightly rounded anteriorly, becoming sharply keeled and more longitudinally elongate posteriorly.
Measurements of holotype (in mm). SVL 343; tail length 601; head length 65.9, width 38.2, depth at eye 28.5; eye–naris distance 22.2; naris–snout distance 11.6; rictus–rictus scales 52; scales around tail base 91; scales around tail 1/3 distance from base 47; mid-body scales 131; gular scales 70; ventrals from gular fold to hind limb insertion 83; total ventral scales 153; dorsals from tympanum to gular fold 25; dorsals from gular fold to hind limb insertion 77; total dorsals 102; supralabials 52; scales around the neck anterior to gular fold 67; and enlarged supraoculars 7L/6R.
Coloration of holotype in preservative. Dorsal surfaces of body, head and limbs black; head nearly uniform black, with dark brown accents dorsal to narial openings and with white pineal scale; trunk with five transverse rows of white blotches; tail with nine irregular bands comprised of diffuse white speckling; forelimbs with sparse white speckling, becoming more regular ventrally; hindlimbs more speckled than forelimbs, and with more speckling ventrally; dorsal surfaces of manus and pes with few white speckles, but surface of pes with slightly more; terminal scale on digits with conspicuous white spot.
Lateral coloration of head consisting of vague, anterior dark brown band extending ventrally from dark brown dorsal markings to narial openings; post-orbitally, lateral surface with longitudinal white stripe extending from the posterior margin of ocular to tympanum ( Fig. 9 View FIGURE 9 ); lateral coloration of the nuchal region consists of bright white mottling, gradually increasing in dorsal extent posteriorly, and terminating at forelimb insertion; lateral portions of trunk with variable white blotches and speckling, with greatest concentration directly posterior to forelimbs, the distribution of which does not correspond to dorsal banding; terminal half of tail solid black dorsally.
Ventral coloration predominately yellow-white throughout; gular region with characteristic dark blotches, becoming more prominent posteriorly; anterior portions of gular region with three triangular projections extending medially; scapular region with irregular dark blotches; trunk with irregular dark triangular projections extending medially from margins, and generally not connecting with projections from opposite side; forelimbs and hindlimbs almost uniform white at insertions, but with increasing dark mottling laterally and in distal portions; manus and pes dark gray; venter of tail similar to that of trunk, but with triangular projections connecting and forming vague bands posteriorly; terminal third of tail solid black ventrally.
Variation. Our small type series exhibits some color variation but specimens are generally phenotypically similar to one another. Four specimens from Mindoro, including the holotype (KU 308437, 335742–44) generally appear darker dorsally, owing to a combination of darker interstitial skin between the scales and fewer whitecolored scales. The holotype (KU 335744) and an additional specimen from Mindoro (KU 308437) were the largest intact specimens examined, and both exhibit a reduction in the size and vibrancy of dorsal banding, with the latter individual having bands that are only faintly discernable. Juvenile specimens from both Mindoro (KU 335742, 335743) and Semirara (KU 305151, 305161, 305163) exhibit much more distinct dorsal bands comprised of series of yellowish to white spots or oscelli. One juvenile from Mindoro ( PNM 9799; formerly KU 308403) appears to be an intermediate, with dorsal bands discernable posteriorly on the trunk and becoming less defined anteriorly. One specimen from Semirara (KU 305161) exhibits a faded reticulate pattern interspersed among the more well-defined dorsal bands, owing to clusters of grayish spots comprised of lightly colored scales and interstitial space. Specimens from Semirara (KU 305151, 305161, 305163) and one from Mindoro (KU 335742) exhibit a more well-defined light-colored longitudinal stripe laterally in the nuchal region, which extends from the ventral margin of the tympanum dorso-posteriorly over the forelimb insertions, joining with the first dorsal trunk band to form a vague “U” shape when viewed from dorsal perspective. Dorsal nuchal coloration varies from uniform black in the holotype (KU 335774) and three paratypes ( PNM 9799, KU 308437, 335772), the presence of 4–10 variable spots or blotches (KU 305151, 308161, 335743), to a more mottled and irregular combination of stipes and blotches posteriorly (KU 305163). One specimen (KU 308437) exhibits a reduction in white speckling on the dorsal surfaces of the limbs.
Ventral coloration is moderately variable. The characteristic gular blotches or spots are present in all specimens, though the number and relative size of blotches is somewhat reduced in four specimens (KU 305161, 308403, 335742, 335743). Two specimens (KU 305151, 305163) exhibit much larger and more conspicuous gular spots. Ventral trunk coloration is less variable, with only three specimens (KU 305161, 308403, 308437) exhibiting reduced transverse dark bands, which generally fail to converge medially. Ventral coloration of the limbs is nearly identical among specimens, with only slight variation in the appearance of black color encroaching on the lateral margins of both fore- and hindlimbs, but generally appearing as partial transverse bands. The holotype (KU 335774) and three paratypes (KU 308437, 305151, 305161) exhibit a more irregular pattern on the margins of the ventral limb surfaces, with dark coloration appearing as an almost reticulate pattern. Ventral coloration of the tail is nearly uniform across specimens, with only minor variation in the extent of medial extension of dark bands. In all specimens, dark bands fail to coalesce anteriorly, but posteriorly to the terminus form a distinct alternating black and white pattern (see Figures 6 View FIGURE 6 and 7 View FIGURE 7 for additional information on color pattern variation).
Coloration in life. Based on the only available image, KU 335742 ( Fig. 8 View FIGURE 8 B), and field notes of the authors. Dorsal ground coloration black, with transverse crossbands of irregularly shaped creamy white to light yellow ocelli from the posterior margin of the nuchal region through the hind limb insertion. Trunk with few, larger ocelli with darkened, ground-colored centers. Additionally, dorso-lateral margins of trunk with smaller, variably distributed light yellow spots or blotches. Lateral margins of head and nuchal region with conspicuous light yellow stripe, extending from the the posterior margin of the ocular, through the tympanum, and midway into the anterior portion of the nuchal region. An additional light yellow stripe extends dorsally and posteriorly from the ventrolateral margin of the nuchal region, and extends to the dorsal region above the forelimb insertion. Snout with distinct and conspicuous bands extending ventrally from dorsal surface.
Dorsal limb surfaces colored similarly to body, but with ocelli irregularly arranged and significantly smaller, lacking any dark-colored center. Forelimbs with larger ocelli dorso-medially.
Ventral coloration creamy white to light yellow, with scattered black pigmentation and with ocelli of ventrolateral surfaces irregularly arranged and significantly smaller, lacking any dark-colored center ( Fig. 8 View FIGURE 8 b).
Ecology, Distribution, and Natural History.—As with all members of the Varanus salvator Complex, V. bangonorum should be considered a habitat generalist, often thriving in a range of habitats from primary forest to more urbanized areas. Like V. dalubhasa , no detailed studies have been undertaken on this taxon specifically, but the findings of Gaulke (in Luxmoore & Groombridge 1989, 1992) are likely relevant. This taxon is most likely to be found in mangrove and riparian habitats (as their common name might suggest). Additionally, most water monitors seek shelter in rock crevices or even trees, often near water sources.
Not surprisingly, Varanus bangonorum has been found on the islands of Mindoro and Semirara ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), both of which are separated by deep water channels that likely have facilitated the isolation of this species and its evolution as a distinct lineage, even during inter-glacial cycles and lowered sea levels ( Voris 2000; Brown & Diesmos 2009).
Again, the findings of Gaulke (1992) are relevent as far as dietary preferences are concerned, and it can reasonably be assumed that the diet of Varanus bangonorum consists primarily of invertebrates and smaller vertebrates (fish, rodents, frogs, snakes). Additionally, as with any species of water monitor, this species is attracted to a number of carion items, with a particular affinity for fish (LJW pers. obs.).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.