
Gromphas dichroa Blanchard, 1846
|
publication ID |
https://doi.org/10.11646/zootaxa.3722.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:20D125E7-8CC0-4942-8AF9-75EA31EBBD53 |
|
DOI |
https://doi.org/10.5281/zenodo.5668593 |
|
persistent identifier |
https://treatment.plazi.org/id/707787C6-596E-6253-FF0A-26D936089CE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Gromphas dichroa Blanchard, 1846 |
| status |
|
3. Gromphas dichroa Blanchard, 1846 View in CoL
Figs. 14 View FIGURES 13 – 15. 13 , 19, 26, 33, 36, 42, 44, 47, 49, 54, 61, 66
Gromphas dichroa Blanchard, 1846: 182 ; Lacordaire 1856: 100; Harold 1869b: 62; Burmeister 1874: 130; Tremoleras 1910: 25; Gillet 1911: 80; d’Olsoufieff 1924: 21, 60, 139, plate IX; Pessoa & Lane 1941: 473; Blackwelder 1944: 208; Martínez 1959: 95; Barattini & Saenz, 1960: 22, 1964: 173; Figueroa et al. 2012: 2.
Bolbites dichrous: Harold, 1869a: 1016 .
Type specimen: Holotype: Not located at MNHN; unknown to us.
Type locality: “Montevideo, près de la mer” (Blanchard 1846).
Redescription: Color: Two general patterns with intermediates ( Fig. 14 View FIGURES 13 – 15. 13 b): 1) Anterior region of clypeus black; remainder of dorsum of head with strong metallic sheen of red, green, and yellow. Pronotum metallic blue on sides and along posterior margin and dark red in center; a narrow strip of metallic yellow and green surrounds entire red area. Metasternum, elytra, and pygidium blue or blue with green reflections ( Figs. 14 View FIGURES 13 – 15. 13 a, 19). 2) Anterior region of clypeus black; remainder of dorsum, metasternum and pygidium predominantly bright light green ( Fig. 14 View FIGURES 13 – 15. 13 c).
Head: Clypeus with truncate apex; genae with very dense and coarse granulation, including region adjacent to eyes ( Fig. 26 View FIGURES 24 – 28 ). Cephalic projection only slightly elevated and with truncate apex ( Fig. 33 View FIGURES 29 – 33 ). Frons granulate at least adjacent to eyes ( Fig. 26 View FIGURES 24 – 28 ).
Thorax: Pronotum globular; lateral region with very dense and coarse granulation (Fig. 19), density of granulation decreasing posteromedially; posteromedian region smooth or with strongly effaced granulation, except for a very narrow strip of dense granulation along posterior margin; posterior fossae present and granulate; posterior margin rounded.
Mesosternum glabrous or with very sparse setae ( Fig. 49 View FIGURES 49 – 52 ). Metasternum clearly punctate at center. Anteromedian angle strongly projecting and narrowed, with truncate apex ( Fig. 49 View FIGURES 49 – 52 ); metasternal area in front of anteromedian angle entirely glabrous (Figs. 19, 49).
Legs: Protibiae very narrow in both sexes; in ventral view, longitudinal carina with a row of tubercles on basal half in males (Figs. 20d, 23b) and simple in females ( Fig. 23 View FIGURES 21 – 23 a). Protibial spur with apex strongly expanded and curved downward ( Figs. 23 View FIGURES 21 – 23 , 45 View FIGURES 34 – 48. 34 ); inner apical angle of protibia with tuft of setae longer in males than in females ( Figs. 23 View FIGURES 21 – 23 a–b). Metatibia very narrow ( Fig. 42 View FIGURES 34 – 48. 34 ). Mesotarsi and metatarsi not enlarged and with apical tarsomeres strongly curved ( Fig. 47 View FIGURES 34 – 48. 34 ). Metatibial spur with apex distinctly curved ( Fig. 39 View FIGURES 34 – 48. 34 ).
Elytra: Striae very fine, simple and effaced, almost imperceptible, especially eighth stria. Sutural margin smooth, distinctly depressed at base.
Abdomen: Pygidium with a pair of weak impressions on sides; basal margin complete and never interrupted by groove of propygidium ( Fig. 35 View FIGURES 34 – 48. 34 ). Sixth sternite densely punctate, remaining sternites smooth.
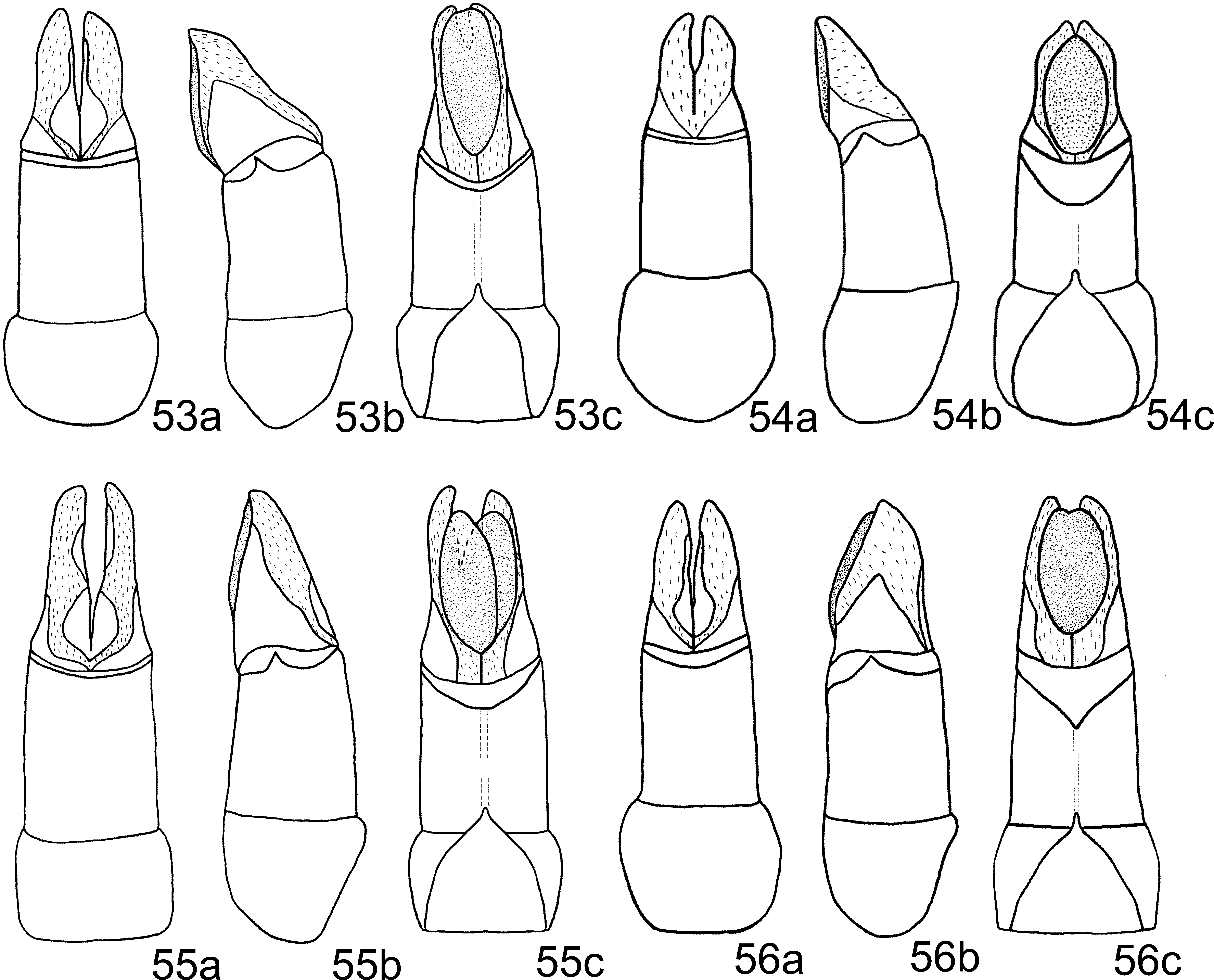
Aedeagus: Phallobase, in ventral view, with wide membranous apical area ( Fig. 54 View FIGURES 53 – 56 c). Medial sclerite very thin and curved ( Fig. 61 View FIGURES 57 – 62 ).
Measurements: Males ( 2 specimens): TL: AV: 12.85; MX: 13; MN: 12.7; SD: 0.21. PL: AV: 12.85; MX: 13; MN: 12.7; SD: 0.21. PW: AV: 7.6; MX: 8.1; MN: 7.1; SD: 0.71. Females ( 5 specimens): TL: AV: 14.84; MX: 16.6; MN: 12.5; SD: 1.54. PL: AV: 12.44; MX: 13.3; MN: 11.8; SD: 0.74. PW: AV: 8.8; MX: 9.7; MN: 8; SD: 0.79. Total ( 7 specimens): TL: AV: 14.27; SD: 1.59. PL: AV: 11.94; SD: 1.1. PW: AV: 8.46; SD: 0.92.
Intraspecific variation and taxonomic discussion: The most striking intraspecific variation in G. d i ch ro a is the color: two distinct patterns are observable, one where green is predominant ( Fig. 14 View FIGURES 13 – 15. 13 c) and the other with dark blue elytra and reddish-blue pronotum ( Fig. 14 View FIGURES 13 – 15. 13 a). However, the specimen from Porto Alegre (Rio Grande do Sul, Brazil) has an intermediate coloration: the elytra are light blue and the pronotum and head are green with a large iridescent yellowish-red spot ( Fig. 14 View FIGURES 13 – 15. 13 b). The green female collected in Santo Tomé (Corrientes, Argentina) is labeled “ Gromphas dichroa var. fulgida nov. A. Martínez det. 1947”, a name never published. There is no reason for the recognition of a separate taxon for specimens of this color pattern, considering that this characteristic is not absolute, as is well demonstrated by the specimen from Porto Alegre, and there is no morphological or distributional difference from the reddish-blue specimens.
Gromphas dichroa has the highest number of unique characters in the genus: (a) truncate apex of clypeus ( Fig. 26 View FIGURES 24 – 28 ); (b) clypeus not folded upwards to any degree (in G. inermis and G. amazonica , the clypeus is strongly folded, while in G. aeruginosa and G. lemoinei it is only slightly); (c) truncate cephalic projection ( Fig. 33 View FIGURES 29 – 33 ); (d) posterior margin of pronotum with narrow margin of granulation, including posterior fossae; (e) mesosternum glabrous or with only very short and sparse pubescence, metasternum glabrous anteriorly to the anteromedian angle (Figs. 19, 49); (f) metatibia very narrow ( Fig. 42 View FIGURES 34 – 48. 34 ); (g) apical mesotarsomeres and metatarsomeres strongly curved ( Fig. 47 View FIGURES 34 – 48. 34 ); (h) elytral suture depressed at base ( Figs. 14 View FIGURES 13 – 15. 13 a–c); and (i) base of pygidium completely margined ( Fig. 35 View FIGURES 34 – 48. 34 ). In addition, (j) the anteromedian angle of metasternum of G. dichroa differs from the other species: it is convex, as in G. amazonica and G. inermis ( Figs. 50–51 View FIGURES 49 – 52 ), but is still much more projected and has a truncate apex ( Fig. 49 View FIGURES 49 – 52 ); in G. lemoinei and G. aeruginosa , the angle is always very flat ( Fig. 52 View FIGURES 49 – 52 ). The smooth sutural margin of elytra (k) is also unique to G. dichroa (in G. aeruginosa the punctation is very sparse, but still present). The color pattern described above for this species (l) is also unique among Gromphas . Finally, the general shape of the medial sclerite of G. dichroa (m) is distinct from that of all other species of Gromphas ( Fig. 61 View FIGURES 57 – 62 ).
Gromphas dichroa shares with G. lemoinei and G. aeruginosa a clypeus with rounded sides ( Fig. 26 View FIGURES 24 – 28 ), and only with G. aeruginosa the presence of posterior pronotal fossae in specimens of any size (in G. lemoinei , these fossae are absent or greatly reduced in larger specimens) and the two tenuous impressions on the sides of pygidium (this feature is very subtle and often absent in G. aeruginosa ). On the other hand, it resembles G. inermis and G. amazonica by the absence of any kind of pronotal prominence; by the protibial spur expanded at the apex ( Figs. 21 View FIGURES 21 – 23 , 45 View FIGURES 34 – 48. 34 ); and by the sexual dimorphism in the tuft of setae at the inner apical angle of protibiae (distinctly more developed in males than in females [ Figs. 23 View FIGURES 21 – 23 a–b]) and in the ventral carina of protibiae (with a row of tubercles in males [Figs. 20d, 23b] and simple in females [ Fig. 23 View FIGURES 21 – 23 a]). Only with G. inermis does G. dichroa share simple elytral striae (in G. dichroa , they are very indistinct and almost imperceptible, especially the eighth and ninth striae, while in G. inermis all striae are easily observed); and only with G. amazonica does it share a smooth posteriomedian region of the pronotum ( Figs. 13, 14 View FIGURES 13 – 15. 13 ).
In Harold’s (1869a) catalog, G. dichroa was transferred to the genus Bolbites , but soon thereafter Harold (1869b) returned this species to its original genus. From all the characters discussed in the description of Gromphas and in the diagnosis of Gromphadina, it is clear that G. di ch ro a belongs in Gromphas and not in Bolbites .
Comments: We have not been able to examine the holotype of G. di ch ro a. Blanchard's (1846) description was based on a single specimen that he believed to be a female (“ Nous n’en avons vu qu’une femelle, que M. d’Orbigny a trouvée à Montevideo, près de la mer ”). d’Olsoufieff (1924) studied the holotype and said it was housed in the MNHN collection, but we have not found it there.
Gromphas dichroa and G. lacordairei Blanchard were published respectively on pages 181 and 182 of volume 6, part 2, of Voyage dans l’Amérique méridionale, series published between 1835 and 1847 in 90 livrasions, seven text volumes and two volumes of plates by Alcide d’Orbigny (Evenhuis 1997). In the sixth volume, Auguste Gaspard Brullé was responsible for the species published on pages 1–60, while Émile Blanchard was for those on pages 61–222. Thus, Blanchard should be regarded as the author of these two species of Gromphas . The publication date usually cited for these species, i.e., 1843, is incorrect. According Sherborn & Woodward (1901) and Evenhuis (1997), livrasion 80, containing pages 169–184 of the sixth volume, was published in 1846.
Martinez (1959) reported G. dichroa from Paraguay. Although we do not know specimens from this country, we examined specimens collected in two Argentine provinces bordering Paraguay, Misiones and Corrientes. It is thus possible that this species is actually present in Paraguay.
None of the females of G. dichroa observed in this study had intact protarsi, all were broken and had only three tarsomeres or less or had the apex of apical tarsomere broken off, which made it impossible to observe its actual morphology ( e.g., if the spiniform projection seen in G. amazonica is present or not).
Bionomics: There are no bionomic studies published on G. dichroa . The specimens examined in this study were collected between October and January.
It is probable that G. dichroa has always been a rare species, but it is interesting to note that all the six specimens known to us with collection date available were collected between the years 1922 and 1947. It is truly remarkable that no specimen has been found over the past 65 years since G. dichroa is distributed in one of the most intensely collected regions of South America. As nothing is known about the natural history of this species, it is difficult to surmise the reasons for the putative disappearance of G. dichroa , but it is possible that the intense human pressure in the Southern Cone (which includes southern Brazil, Paraguay, Uruguay, Argentina, and Chile) has acted negatively on what were already small populations.
Geographic distribution: Chacoan subregion: Chaco and Pampa. BRAZIL: Rio Grande do Sul: Cerro Largo, Porto Alegre. PARAGUAY. ARGENTINA: Misiones: Concepción (Santa Maria). Corrientes: Santo Tomé. Buenos Aires. URUGUAY: Tacuarembó, Soriano, Flórida, Montevidéu. ( Fig. 66 View FIGURE 66 ).
Material examined: 3 males and 6 females. ARGENTINA: BUENOS AIRES: Anchorena, X.1935, M. J. Viana col.— 1 male (CMNC). CORRIENTES: Santo Tomé, XII.1922, without collector— 1 female (CMNC). MISIONES: Concepción, Santa Maria, XI.1947, M. J. Viana col.— 1 male (MLPA). BRAZIL: RIO GRANDE DO SUL: Cerro Largo (former Serro Azul), I.1931 (or 1933?), without collector— 1 male (CEMT); Porto Alegre, X.1928, without collector— 1 female (MZSP). URUGUAY: without date and collector— 3 females (MZSP) and 1 female (CEMT).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Scarabaeinae |
|
Tribe |
Phanaeini |
|
Genus |