
Bryobia ( Bryobia ) watersi Manson, 1967
|
publication ID |
https://doi.org/10.24349/acarologia/20204367 |
|
DOI |
https://doi.org/10.5281/zenodo.4527074 |
|
persistent identifier |
https://treatment.plazi.org/id/750B87C3-2A56-4722-C1C2-F80BFDA2FC7A |
|
treatment provided by |
Felipe |
|
scientific name |
Bryobia ( Bryobia ) watersi Manson, 1967 |
| status |
|
Bryobia ( Bryobia) watersi Manson, 1967
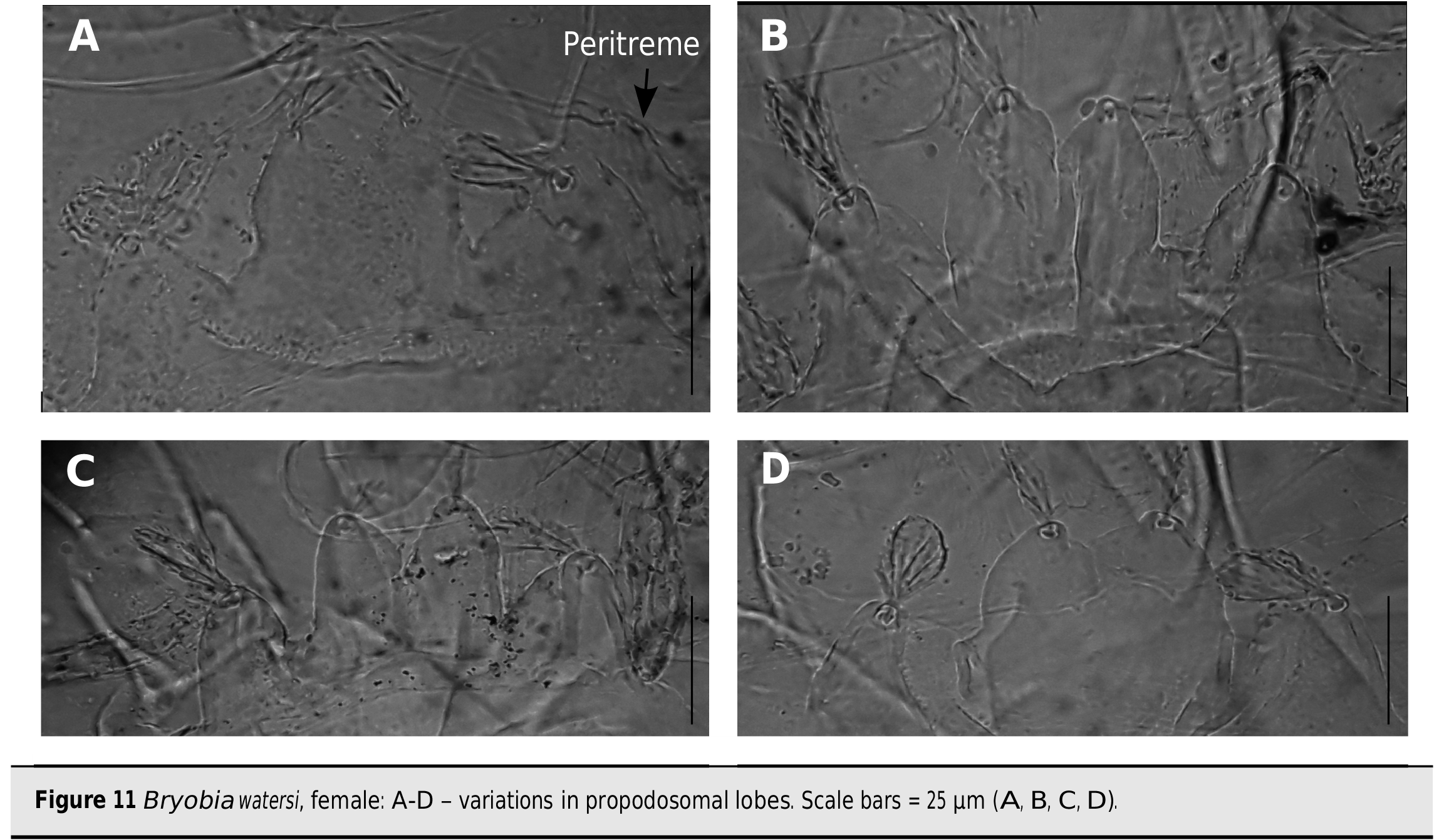
The specimens of this species were identified as Bryobia sp. and misidentified as B. ( B.) graminum and/or B. ( B.) kissophila in previous studies ( Barbar, 2014, 2018). This was due to species identification only based on the morphological characteristics of females and particularly on variations in the shape of propodosomal lobes. In the present study, comparisons of all the specimens collected (including females, a male and juveniles, Figures 11-15 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 ) with the detailed redescriptions of B. ( B.) graminum , B. ( B.) kissophila and B. ( B.) praetiosa by Mathys (1957) and the original description of B. ( B.) watersi by Manson (1967) were undertaken. This led us to the conclusion that the morphological characteristics of the Syrian specimens fit well with those provided in the original description B. of ( B.) watersi by Manson (1967) even if few morphological small differences were found: the dorsal body setae are slightly longer in Syrian females (there are no obvious differences in juveniles); genu II and tarsus IV of Syrian females with 6 and 14 setae respectively vs. 5 and 15 setae; genu II-III of Syrian male with 6 – 5 setae vs. 4[5] – 6; and tarsus III of Syrian deutonymphs with 10 setae vs. 11[12].
Remarks — This species was collected from several host plants surrounding a citrus orchard located at Al-ya’robiyah, Latakia governorate, Syria ( 35°30’24” N, 35°48’33” E) and was found on: Amaranthus retroflexus L., a new host plant (five females, 15 April 2014, previously identified as Bryobia sp.), Malva sylvestris L. (five females, two deutonymphs, three protonymphs and two larvae, 1 January 2014; four females, 15 February 2014; three females, 7 April 2015; two females, two protonymphs and one larva, 18 February 2016, previously misidentified as B. ( B.) graminum ; six females and one larva, 16 April 2016, previously misidentified as B. ( B.) kissophila ), Urtica urens L., a new host plant (three females and one protonymph, 15 April, 2014, previously identified as Bryobia sp.) and Trifolium sp., a new host plant (four females, three deutonymphs and one male, 14 April, 2015, previously identified as Bryobia sp.).
Among the specimens of B. ( B.) watersi mentioned above, two females collected from A. retroflexus have only three propodosomal lobes each with a single seta ( Figures 15 View Figure 15 A-B). Initially, the specific identification of these specimens led to a result that they are either new specimens (or closely related species) of the two “trilobed” species B. bakeri Zaher et al. (1982) and B. aegyptiacus Zaher et al. (1982) or they are aberrant Bryobia specimens. Actually, the overall propodosomal lobe shapes of the trilobed Syrian specimens are closer to those of B. bakeri ( Figure 15A View Figure 15 ) than those of B. aegyptiacus . Nevertheless, the Syrian specimens differ from those two species by the palptarsus setal count [unusual counts are found in the two species described by Zaher et al. (1982)] and by small differences in the global leg setal counts. These results led to conclude that the Syrian trilobed specimens did not belong B. to bakeri nor to B. aegyptiacus .
Indeed, despite the differences between the propodosomal lobes of the specimen presented in Figure 15B View Figure 15 (typical of abnormal ”asymmetrical” lobes; the outer lobe of the left side is obviously missing) and those of the specimen presented in Figure 15A View Figure 15 (the axis of symmetry passes through the middle of the single inner lobe), we concluded that these females are abnormal individuals of B. ( B.) watersi for several reasons:
(1) Both specimens are morphologically identical to B. ( B.) watersi [i.e. having similar leg ambulacra, leg chaetotaxy and articles dimensions, same shape and length of dorsal body setae and peritreme etc.].
(2) Both specimens were collected together with specimens of B. ( B.) watersi (same host plant, same date and place of collection).
(3) Several attempts carried out in 2015 and 2016 to re-collect additional trilobed individuals (in the same location where they were collected first) were unsuccessful and all recollected individuals were B. ( B.) watersi .
(4) Propodosomal lobe aberrations have already been reported in several Bryobia species ( Arabuli and Auger, 2016; Fashing et al. 2016; Smiley and Baker, 1995).
This variability in the propodosomal lobe shape found in the two Syrian trilobed individuals of B. ( B.) watersi guided us to question about the taxonomical value of the number of propodosomal lobes used to separate B. bakeri and B. aegyptiacus from other Bryobia species. Several arguments tend to show that these specimens could be teratological forms rather than species with a particular propodosomal lobes pattern:
(1) Specimens are rare: like the Syrian trilobed specimen of B. ( B.) watersi of the Figure 15A View Figure 15 , only one specimen of B. bakeri and one of B. aegyptiacus are known. Although Smiley and Baker (1995) reported a possible additional female of B. bakeri , it could belong to another species because its leg setal count is far different from that of the type specimen of B. bakeri (it shares the same setal count only on five leg articles; as a comparison, the Syrian trilobed B. ( B.) watersi are closer to B. bakeri for the reason that they share the same setal count on 12 leg articles).
(2) Specimens with three propodosomal lobes (each bearing one seta) are known to occur in several species of Bryobia: Smiley and Baker (1995) mentioned that in a few species of Bryobia some aberrant females (with two or three propodosomal lobes) appear sometimes. Since that work, several cases of Bryobia species with three propodosomal lobes have been reported ( Arabuli and Auger, 2016; Fashing et al. 2016). In the detailed study by Fashing et al. 2016, it was demonstrated that both morphotypes (with three or four propodosomal lobes) of Bryobia abyssiniae Fashing and Ueckermann, 2016 belong to the same species, and about 9.5% of observed specimens had a single propodosomal inner lobe (with a single seta v 1).
(3) The two Syrians trilobed specimens are conspecific despite the fact that one of them is obviously an aberrant form (asymmetry) and the other has a symmetrical propodosomal lobe pattern similar to that found in B. bakeri .
This tends to show that a bryobiine mite with an unpaired inner propodosomal lobe bearing a unique seta v 1, can be an aberrant specimen despite a symmetrical propodosomal trilobed lobe pattern.
In our opinion, all these elements together strongly suggest that B. bakeri and B. aegyptiacus would be more aberrant individuals of two species of Bryobia (four-lobed) than species characterized by unpaired inner propodosomal lobe. Even if the data are insufficient to assign these “trilobed” species to an existing four-lobed Bryobia species, the demonstration presented here is consistent with the synonymy of the genus Septobia with the genus Bryobia by Bolland et al. (1998).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Bryobiinae |
|
Tribe |
Bryobiini |
|
Genus |