
Eulepidina sp.
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2021v43a13 |
|
publication LSID |
urn:lsid:zoobank.org:pub:7EB95293-B048-41D0-9237-F66CBFDD43E5 |
|
DOI |
https://doi.org/10.5281/zenodo.5040973 |
|
persistent identifier |
https://treatment.plazi.org/id/7668E43B-FFCB-FFDB-BA70-4BB2FE93144E |
|
treatment provided by |
Felipe |
|
scientific name |
Eulepidina sp. |
| status |
|
( Fig. 5H, I View FIG )
REMARKS
The identification is based only upon axial sections. The test is lenticular, its diameter ranges from 8.1 to 11.2 mm and its thickness from 1.1 to 1.7 mm. The equatorial chambers are polygonal in shape.
DISCUSSION ON THE LARGER FORAMINIFERAL RECORD
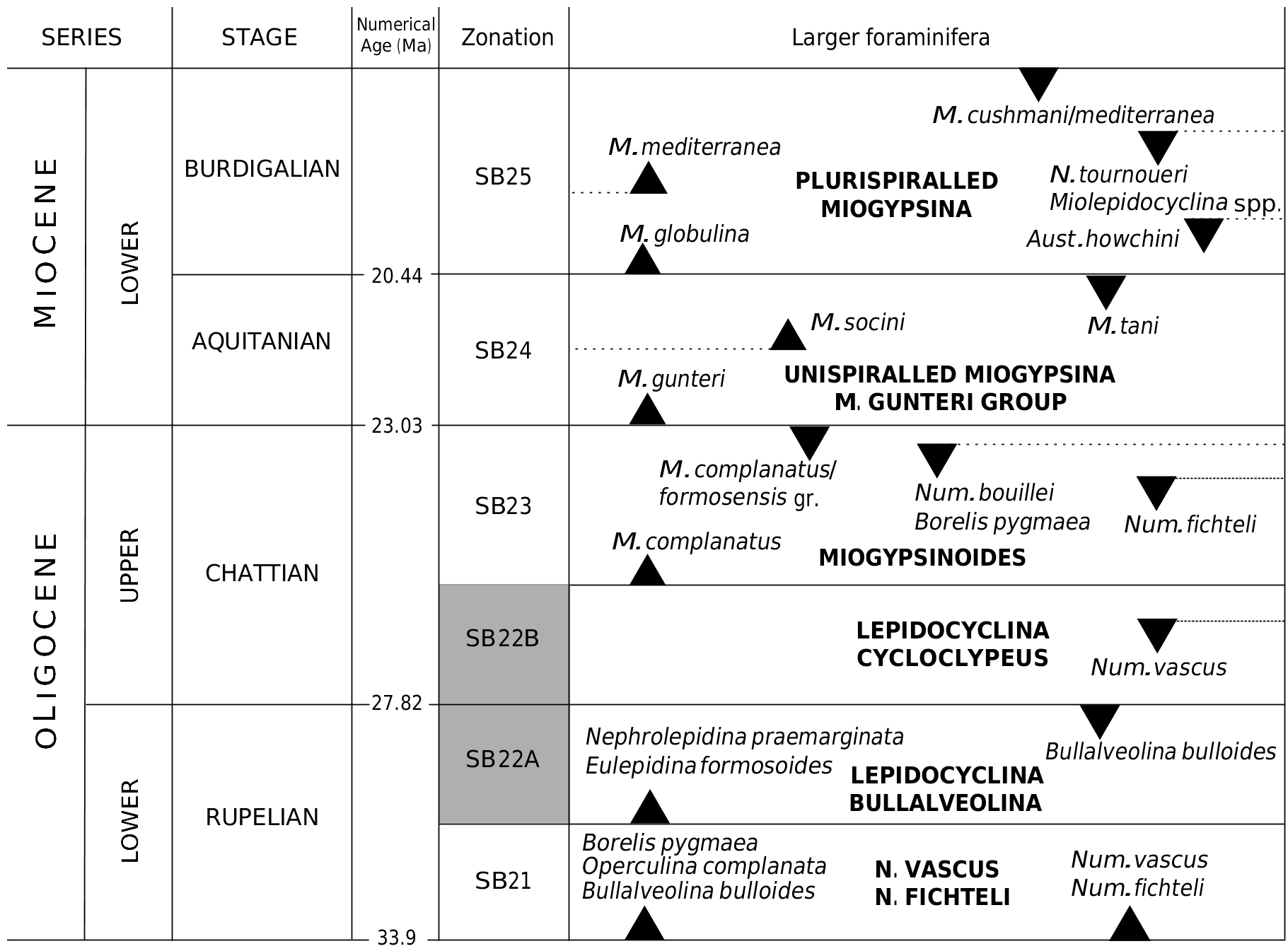
The study of Oligocene hyaline larger foraminifera, based on the biometric study of the nummulitids and lepidocyclinids, from western Taurids, gives insights on their palaeobiogeography in the Tethys. These groups show that they are closely related with the coeval assemblages known from the European basins. The larger benthic foraminiferal association from the Burdur-2 section correlates with the Shallow Benthic Zone (SB) 22, late Rupelian-early Chattian, based on the time range of Nummulites fichteli and Nephrolepidina praemarginata ( Cahuzac & Poignant 1997) . In the upper part of the Burdur-2 section, N. praemarginata and N. fichteli are associated with Operculina complanata , Eulepidina sp. , Asterigerina sp. , Planorbulina sp. , and Amp- histegina sp. ( Fig. 6 View FIG ). Some species of this assemblage have a longer stratigraphical range such as Operculina complanata which extends from the base of the Oligocene to the Late Miocene ( Cahuzac & Poignant 1997). This evidence dates
the Hayrettin Formation as late Rupelian-early Chattian (SB 22).
In the Mediterranean realm, the evolutionary lineage of Nephrolepidina praemarginata - N. morgani - N. tournoueri is used as key criteria in Oligo-Miocene biostratigraphy. Although Douvillé (1908, 1925) reported N. praemarginata from the early Oligocene of Italy and from the Oligocene-Miocene transition in France, Cahuzac & Poignant (1997) demonstrated that this form is limited to the biozone SB 22.
This taxon is poorly documented in Turkey. Some specimens of this species have been described in eastern Turkey
Numerical SERIES STAGE Age (Ma) Zonation Larger foraminifera M. cushmani / mediterranea M. mediterranea E BURDIGALIAN SB25 N. tournoueri N PLURISPIRALLED MIOGYPSINA Miolepidocyclina spp. E C LOWER M. globulina Aust. howchini O 20.44 I M M. socini M. tani AQUITANIAN SB24 M. gunteri UNISPIRALLED MIOGYPSINA M. GUNTERI GROUP 23.03 M. complanatus / formosensis gr. Num. bouillei SB23 Borelis pygmaea Num. fichteli E M. complanatus N UPPER MIOGYPSINOIDES CHATTIAN E C SB22B LEPIDOCYCLINA O CYCLOCLYPEUS Num. vascus G I L 27.82 O Nephrolepidina praemarginata SB22A Bullalveolina bulloides Eulepidina formosoides LEPIDOCYCLINA BULLALVEOLINA LOWER RUPELIAN Borelis pygmaea Operculina complanata N. VASCUS Num. vascus SB21 Bullalveolina bulloides Num. fichteli N. FICHTELI 33.9
as Nephrolepidina partita based on the presence of coarse granules in the umbonal part of the test ( Sirel 2003; Gedik 2014, 2015). According to Sirel (2003), N. praemarginata is associated in the Sarıbuğday section (Elazıǧ region, eastern Turkey) with Nummulites fichteli , N. vascus , N. cf. germanicus, Borelis pygmaea , B. inflata , B. merici , Operculina complanata , Neorotalia lithothamnica , N. pinarensis , N. cf. tectoria, N. viennoti , Austrotrillina asmariensis , Eulepidina sp. , Thalmannita sp. and other smaller benthic foraminifera. This foraminiferal association corresponds with the biozones SB 21 and SB 22 of Cahuzac & Poignant (1997). It is present at the Rupelian/Chattian boundary (G. opima opima zone; P 21) in the Kelereşdere section (Muş region, eastern Turkey) which yielded N. partita , B. merici , B. cf. inflata , A. striata , Eulepidina sp. corresponding to the biozone SB 22.
According to Özcan et al. (2009), in the Korkuteli area (namely Kabaktepe, Yuvaköy, Hacıbekar sections) and in the Tavas-Burdur Basin (Kızılcaağaç section), N. praemarginata is associated with Eulepidina dilatata , Operculina complanata , Nummulites kecskemetii , Heterostegina assilinoides . This foraminiferal association corresponds to SB 22B (early Chattian).
As inferred by Gedik (2014, 2015), N. praemarginata is associated in the Develi section (W of Malatya, eastern Turkey) with Archaias kirkukensis , Austrotrillina asmarien- sis, A. brunni , Peneroplis cf. laevigatus , Nephrolepidina sp. This foraminiferal association corresponds with biozones SB 21, 22 of Cahuzac & Poignant (1997). It occurs at the Rupelian/Chattian boundary in the Edilme section (W Malatya, eastern Turkey) together with Heterostegina assilinoides , Nummulites cf. vascus , Eulepidina cf. formosoides , Risananeiza crassaparies , and Neorotalia lithothamnica corresponding to the biozone SB 22.
An early Rupelian to early Chattian (SB 21-22B) age is known for Nummulites fichteli in Europe and Mediterranean area ( Cahuzac & Poignant 1997; Sirel 2003; Gedik 2008). This species is known from the lower Oligocene shallow marine deposits in central and southern Iran (SB 21-22A) ( Ehrenberg et al. 2007; Van Buchem et al. 2010; Yazdi- Moghadam 2011). In the Denizli Basin, which is located adjacent to the study area, Nummulites fichteli is associated with Nummulites vascus and Operculina complanata , indicating a Rupelian-early Chattian age (SB 21-22; Gedik 2008). Nummulites vascus is a key species for the Oligocene but we did not find it in the examined strata.
CONCLUDING REMARKS
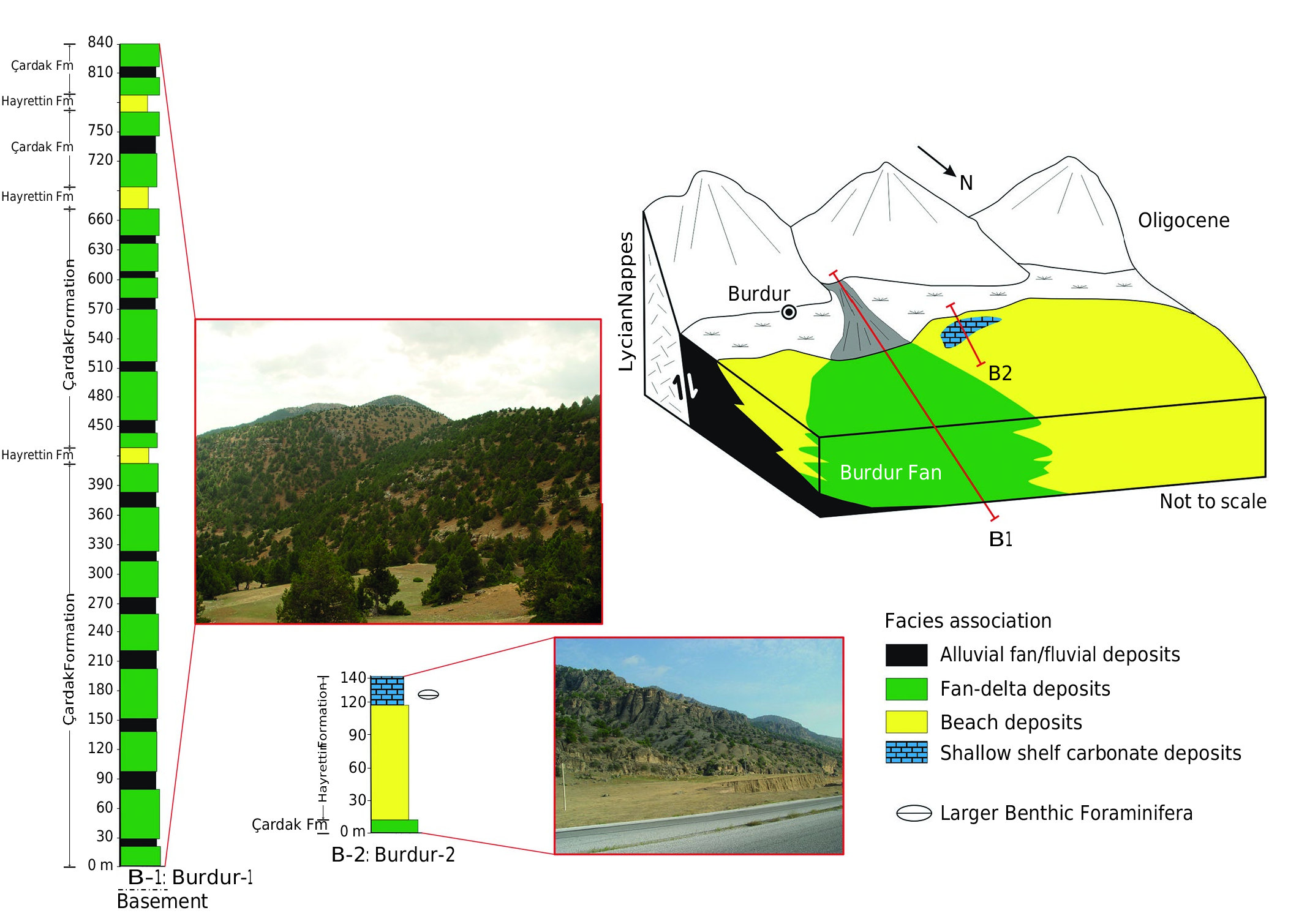
Southwestern Anatolia has been under the influence of an extension regime since the Oligocene ( Seyitoğlu et al. 2004; Karadenizli et al. 2009, 2017), with thick sequences deposited in graben basins opened by normal faults.An example of this system is seen in the Burdur region where a 840 m thick fan sequence developed in a graben basin ( Fig. 3 View FIG ). A simple stratigraphic succession developed in the Burdur Fan during the Oligocene, which consists of the Çardak and Hayrettin formations. The ten lithofacies have been grouped into four facies associations ( Fig. 2 View FIG ) based on their lateral and vertical relations ( Fig. 3 View FIG ). Sedimentary environments of these facies associations show that the Burdur region was invaded by a shallow sea from the west, and that the sea penetrated into Anatolia.
The basin fill is controlled by the Burdur Normal Fault which formed the southern edge of the basin. Along this margin, alluvial-fluvial fan and fan-delta facies associations developed, while in the northern part sedimentation was limited to fan-delta successions ( Fig. 3 View FIG ). These facies mainly consist of coarse-grained sediments derived from the Taurus tectonic unit. There is no evidence for sediment derivation from the metamorphics of the Menderes Massif. A beach facies is present towards the center of the basin ( Fig. 3 View FIG ). Ramp-type carbonate platforms also developed depending on the shelf topography in the intervals when clastic input was minimal ( Fig. 3 View FIG ).
A moderately diverse and well-preserved assemblage of hyaline and porcellaneous larger foraminifera was found in the late Rupelian-early Chattian shallow marine deposits of the Hayrettin Formation in the Burdur Basin, SW Anatolia. The foraminiferal association includes 7 genera. Larger foraminifera from the Hayrettin Formation, classified under the genera Nummulites , Operculina , Nephrolepidina , Eulepidina , Planorbulina , Amphistegina and Asterigerina show close similarity to the coeval assemblages already known from the European and circum-Mediterranean marine sedimentary sequences. Their western Tethyan affinity allowed us to apply the standard biozonation scheme (SB zonation) for our study area in SW Turkey. The late Rupelian-early Chattian age, i.e., SB 22 Zone of Cahuzac & Poignant (1997) for the Burdur-2 section is documented by the presence of Nephrolepidina praemarginata , together with Nummulites fichteli . This evidence dates the Hayrettin Formation as late Rupelian-early Chattian (SB 22).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |