
Hamatopeduncularia bagre Hargis, 1955
|
publication ID |
https://doi.org/10.11646/zootaxa.4127.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:D5AEC22D-65C0-49B6-92C5-B8D1BCBBF91F |
|
DOI |
https://doi.org/10.5281/zenodo.5699408 |
|
persistent identifier |
https://treatment.plazi.org/id/7A05A870-3743-FFC4-E3B5-FE0CFD141992 |
|
treatment provided by |
Plazi |
|
scientific name |
Hamatopeduncularia bagre Hargis, 1955 |
| status |
|
Hamatopeduncularia bagre Hargis, 1955 View in CoL
( Figures 37–46 View FIGURES 37 – 40 View FIGURES 41 – 46 )
Syn. Hamatopeduncularia bagrae Hargis, 1955 (misspelling in Tripathi, 1959, page 59) Hargitrema bagre ( Hargis, 1955)
Tripathi 1959; Hargitrema bagrae ( Hargis, 1955) Triparthi 1959 (misspelling in diagnosis). Hamatopeduncularia bagre: Hargis (1955) : 188–189, Figs 34–38 View FIGURES 30 – 36 View FIGURES 37 – 40 (descr); Lim et al. (2001): 174–176 (citat). Hargitrema bagrae: Triparthi (1959) : 60 (diag); Paperna (1977): 112, photo 2 (citat).
Hargitrema bagre: Yamaguti (1963) 69–70, pl. 13, Fig. 120 (citat.).
Type-host: Bagre marinus (Mitchill)
Site of infection: Secondary lamellae of the gills.
Type-locality: Alligator Harbor, Franklin County, Florida, USA.
Other records: Bagre bagre (Linnaeus) , Bombom Village, Municipality of Viseu, State of Pará, Brazil ( 01º11’48”N, 46º08’24” W); B. bagre , Caratateua Village, Municipality of Bragança, State of Pará, Brazil ( 0°59’ N, 46°44’, 46°44’ W); B. bagre , Fishing village of Ajuruteua, Municipality of Bragança, State of Pará, Brazil ( 0°49’31”N, 46°36'29"W); B. bagre , Fish market, Municipality of Bragança, State of Pará, Brazil.
Specimens studied: 1 holotype USNPC 49339; 25 Vouchers, CHIOC 38273–38280, INPA 680–683, MPEG 061–062.
Prevalence of infection: 33 of 74 (45%) of Bagre bagre .
Comparative measurements: Table 6 View TABLE 6 .
Redescription: Based on 21 adult specimens; 10 mounted in Gomori’s trichrome, 11 mounted in Hoyer’ medium. Body fusiform ( Fig. 37 View FIGURES 37 – 40 ), total length excluding haptor 1307 (1010–1560; n=3) long, fusiform, 256 (240– 280; n = 5) wide, usually at level of germarium. Tegument smooth. Cephalic margin tapered; moderately developed terminal lobes; three bilateral pairs of head organs with rod-shaped secretion; cephalic glands unicellular, posterolateral to pharynx. Eyes 4, equidistant; members of posterior pair of eyes larger than anterior pair; accessory granules absent or few in cephalic area, spherical. Mouth subterminal, midventral; pharynx subspherical, 136 (120–170; n = 7) long, 97 (90–115; n=5) wide; oesophagus short; intestinal caeca, nonconfluent, lacking diverticula. Genital pore opening midventral, anterior to copulatory complex; genital atrium muscular. Testis postgermarial; testis dorsal to germarium. Testis elongate fusiform, about 1/3 body length, 400 (350–450; n=2) long, 100 (100–200; n=3) wide. Vas deferens looping left intestinal cecum; seminal vesicle a dilatation of vas deferens; proximal portion of seminal vesicle sigmoid, crossing midline, dorsal to ootype, uterus; distal portion of seminal vesicle dilated, looping before connecting with male copulatory organ. Prostatic reservoir not observed. Copulatory complex comprising male copulatory organ, accessory piece; male copulatory organ, sclerotized, a coiled tube with 1 counterclockwise ring; distal aperture subterminal; base with sclerotized margin. Accessory piece sclerotized, non-articulated with male copulatory organ; proximal portion spoon shaped; distal portion with two branches, small branch with acute termination, large branch with spines at midpoint, rounded termination ( Fig. 39 View FIGURES 37 – 40 ). Germarium 100 (100–200; n=3) long, 50(50–100; n=3) wide, constricted at level of seminal receptacle; proximal portion subovate, distal portion elongate ( Fig. 38 View FIGURES 37 – 40 ). Vagina comprising vaginal aperture with dextroventral opening; vaginal vestibule with soft tissue at proximal portion; distal portion of vaginal vestibule sclerotized, cup shaped; vaginal canal sclerotized, short ( Fig. 40 View FIGURES 37 – 40 ). Seminal receptacle pyriform, distal end ascendant; Mehlis’ glands bilateral to ootype. Vitellaria coextensive with caeca. Egg tetrahedral with short filament. Peduncle elongate. Haptor digitate, (usually 12), 342 (270–430; n = 6) long, 257 (220–290; n = 3) wide. Anchors dissimilar. Ventral anchor ( Fig. 44 View FIGURES 41 – 46 ), three times smaller than dorsal anchors, 60 (52–62; n = 6) long, base 17 (17; n = 4) long, inconspicuous roots, anterior tip covered with sclerotized cap; evenly curved shaft and point. Dorsal anchor ( Fig. 41 View FIGURES 41 – 46 ) 248 (270–315; n = 7) long, base 47 (42–62; n = 7) long, with elongate superficial root 228 (200–250; n=21) long, inconspicuous deep root, both root tips covered with sclerotized cap; evenly curved shaft and point; internal shaft with sclerotized cap shield shaped; external shaft with expansion keel shaped. Haptor with 3 (1 midventral, 2 of Bagre marinus (USNPC 49339) and Bagre bagre . MCO= male copulatory organ. mid-dorsal) haptoral bars. Ventral bar ( Fig. 42 View FIGURES 41 – 46 ), broadly V shaped, with bifurcated ends 104 (92–120; n = 9) long, 7 (7–7; n = 20) wide. Paired dorsal bars with robust muscles connecting both bars, each dorsal bar with 204 (178– 254; n = 10) long, 40 (29– 52; n = 10) wide, ( Figs. 45–46 View FIGURES 41 – 46 ), expanded ends with anterior projection twisted dorsally. Hooks ( Fig. 43 View FIGURES 41 – 46 ) similar, 17 (14–19; n = 16) long, erect thumb, lightly curved shaft, short point; shank divided into two subunits, proximal 1/3 of shank inflated; filamentous hook loop extending to near beginning of shank dilation.
Remarks: Hamatopeduncularia bagre Hargis, 1955 was proposed to accommodate a species from the gills of Bagre marinus collected from Alligator Harbor, Franklin County, Florida, USA. The species was characterized mainly by the possession of hooks on haptoral digits, two articulated dorsal bars, and dissimilarity in the size of anchors. Tripathi (1959) considered that H. bagre did not belong to Hamatopeduncularia , and proposed Hargitrema Tripathi, 1959 to accommodate it as Hargitrema bagrae ( Hargis, 1955) Tripathi, 1959 . The genus was defined as having two pairs of unequal anchors, three bars (one ventral, two dorsal), and intestinal caeca posteriorly united by an isthmus. Yamaguti (1963), Paperna (1977) and Lim (1996) accepted Hargitrema as a valid genus. However, Lim et al. (2001) re-assigned Hamatopeduncularia bagre and considered Hargitrema a junior synonym of Hamatopeduncularia . Comparison of the holotype of H. bagre (USNPC 49339) with specimens collected from the gills of Bagre bagre from Atlantic Amazon coast of Brazil allowed us to detect that Hargis (1955) erroneously identified the overlap of vitelline follicules posteriorly to testis as an isthmus uniting intestinal caeca.
The study of available specimens allowed us to confirm also that the dorsal bar is divided into two pieces, as proposed by Hargis (1955) in the description of H. bagre . Boeger & Kritsky (1997) found 1 ventral, 1 dorsal bars in the haptor to be a synapomorphy for the clade Dactylogyrinea+Tetraonchinea, while 1 ventral, 2 dorsal bars represented a synapomorphy for two independent clades in both suborders: Sundanonchidae + Tetraonchidae , and Diplectanidae + Pseudomurraytrematidae . Occurrences of 1 ventral, 2 dorsal bars in the haptor of some dactylogyrids species ( i.e. species of Curvianchoratus Hanek, Molnar & Fernando, 1974, Trinibaculocauda Tripathi, 1959, Trinibaculum Kritsky, Thatcher & Kayton, 1980, Thaparocleidus tengra ( Tripathi, 1959) Lim, 1996 and Hamatopeduncularia bagre ) is apparently derived within the family and represents autapomorphies for those taxa.
Except for the presence of double dorsal bar, Hamatopeduncularia bagre shares features typically associated with the genus ( i.e., haptoral digits, absence of expanded deep roots on the ventral anchors), which supports the maintenance of this species in Hamatopeduncularia .
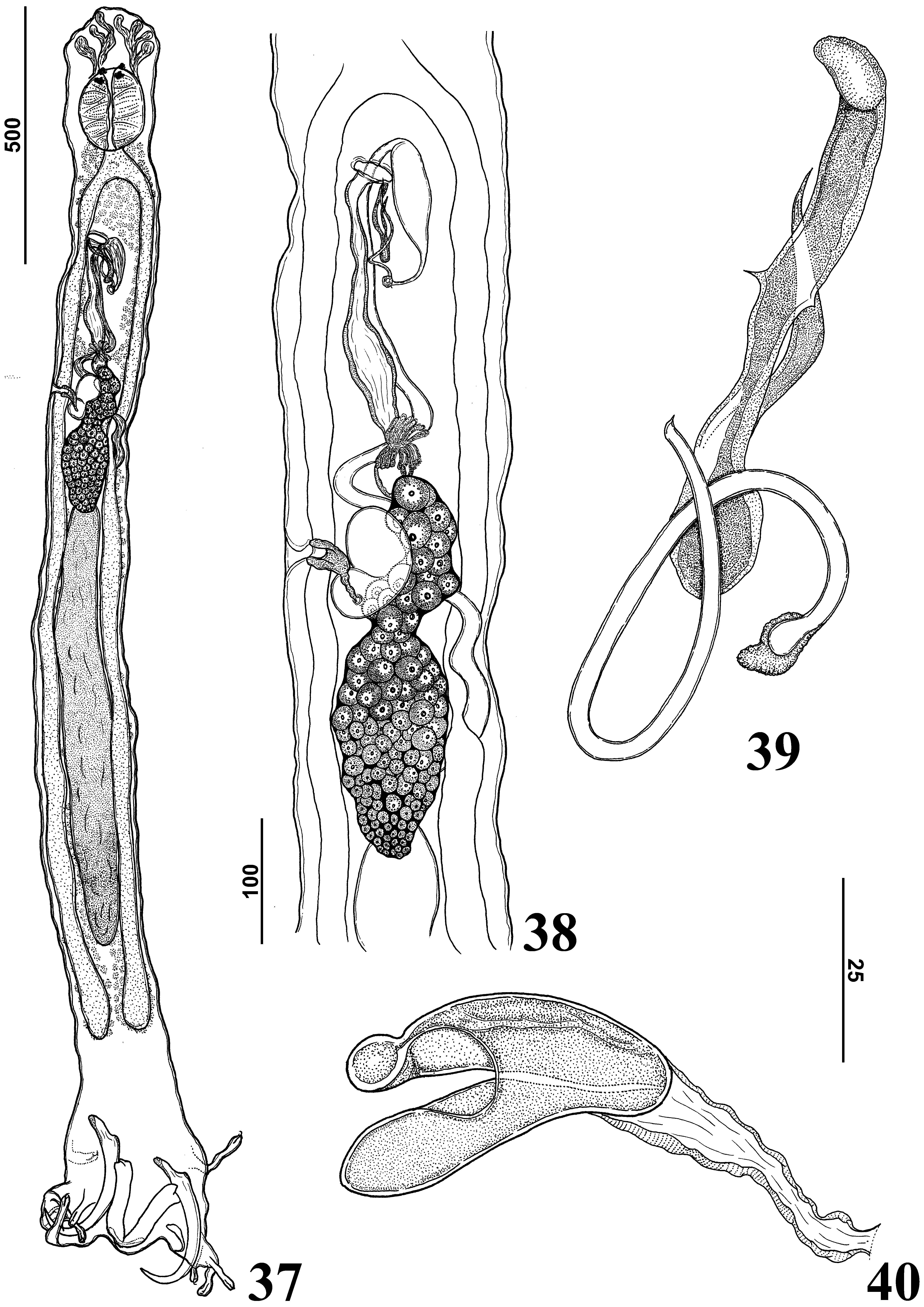
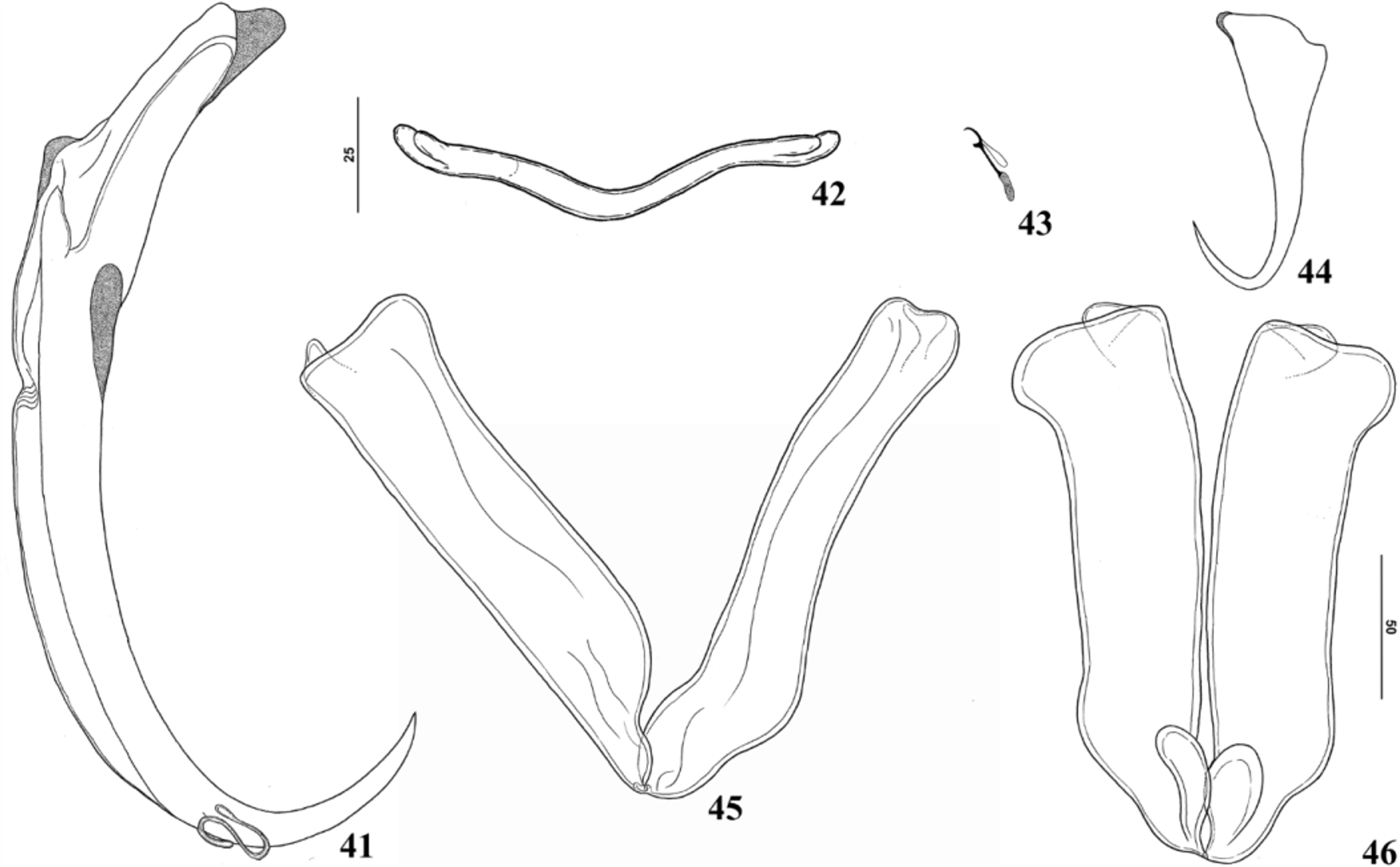

TABLE 6. Comparative measurements (in µm) of specimens of Hamatopeduncularia bagre Hargis, 1955 from the gills
| B. bagre | N | B. marinus | N |
|---|---|---|---|
| Body | |||
| Length 1307 (1010–1560) | 3 | 1427 | 1 |
| Width 256 (240–280) | 5 | 238 | 1 |
| Haptor | |||
| Length – | – | – | – |
| Width – | – | – | – |
| Pharynx | |||
| Length 136 (120–170) | 7 | 92 | 1 |
| Width 97 (90–115) | 5 | 64 | 1 |
| MCO length – | – | – | – |
| MCO ring – | – | – | – |
| Ventral Bar | |||
| Length 45 (35–65) | 9 | – | – |
| Width 207 (183–256) | 9 | – | – |
| Dorsal Bar | |||
| Length 5 (5–6) | 4 | – | – |
| Width 94 (68–112) | 4 | – | – |
| Ventral Anchor | |||
| Inner 51 (50–52) | 4 | – | – |
| Outer 60 (52–62) | 6 | 49) | 1 |
| Base 17 (17–17) | 4 | 14 | 1 |
| Dorsal Anchor | |||
| Inner 284 (270–315) | 7 | 199 | 1 |
| Outer 228 (200–250) | 21 | – | – |
| Base 47 (42–62) | 7 | 12 | 1 |
| Hooks 17 (14–19) | 16 | – | – |
| Germarium | |||
| Length – | – | – | – |
| Width – | – | – | – |
| Testis | |||
| Length – | – | – | – |
| Width – | – | – | – |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hamatopeduncularia bagre Hargis, 1955
| Domingues, Marcus V., Soares, Geusivam B. & Watanabe, Alana 2016 |
Hargitrema bagre:
| Yamaguti 1963 |