
Anelosimus, Simon, 1891
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2004.00120.x |
|
persistent identifier |
https://treatment.plazi.org/id/7E1687E1-4228-6C1D-FC7D-FAF38881FD02 |
|
treatment provided by |
Diego |
|
scientific name |
Anelosimus |
| status |
|
ANELOSIMUS View in CoL View at ENA : CLADE 24
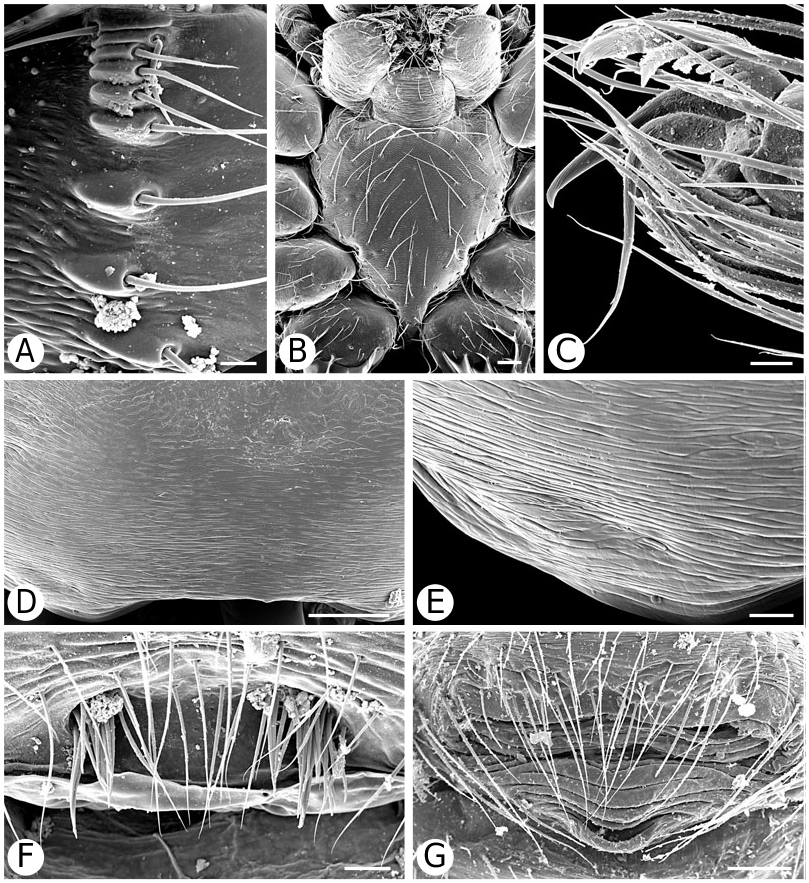
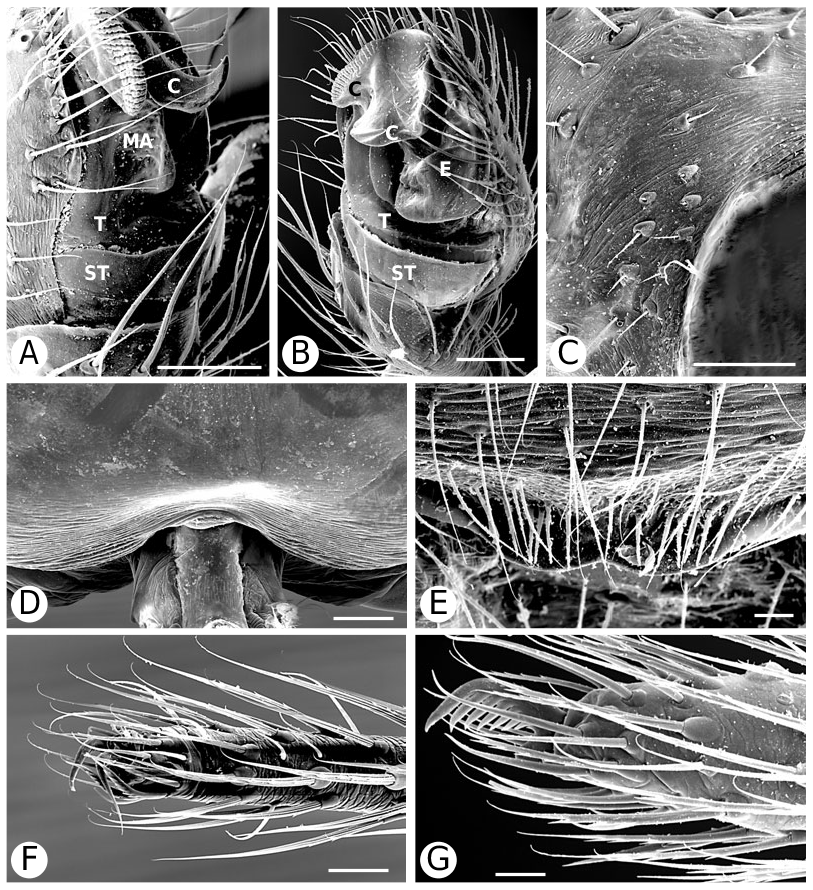
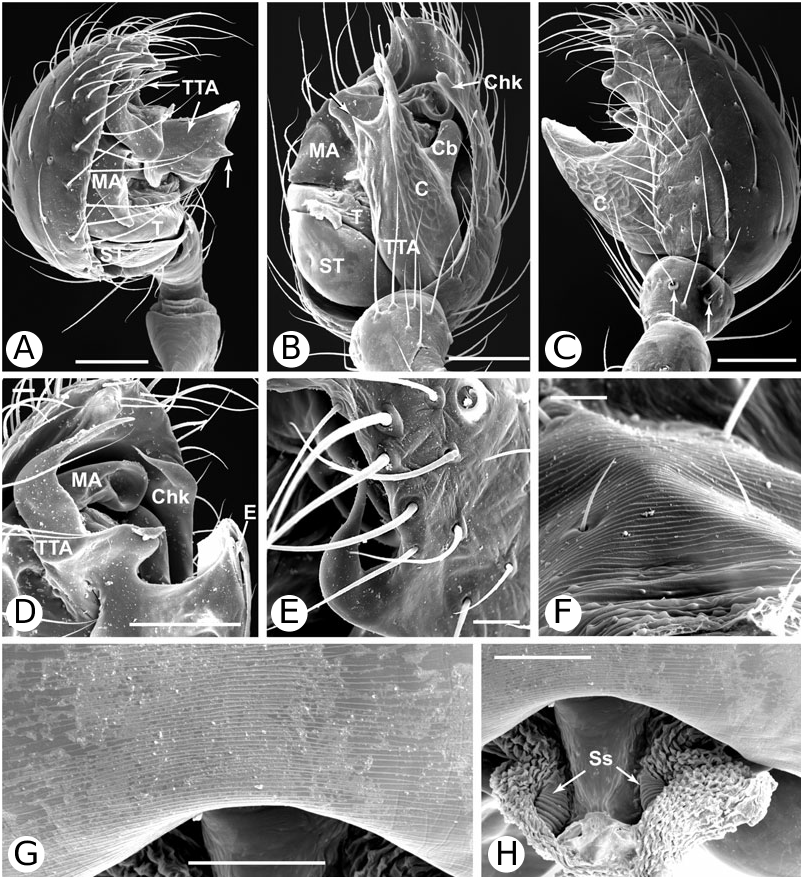
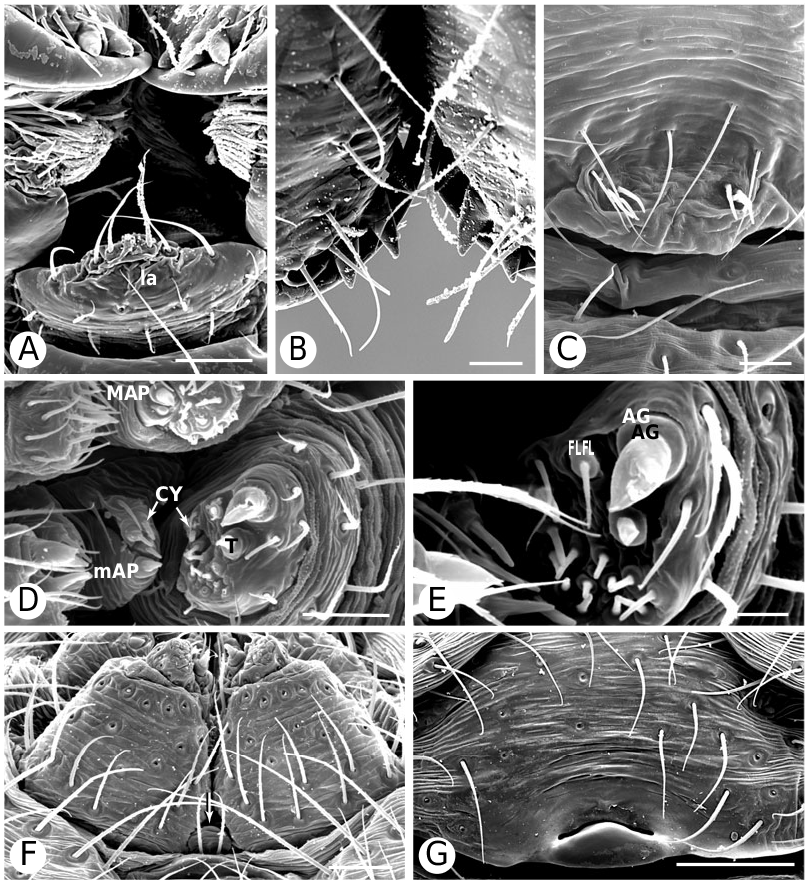
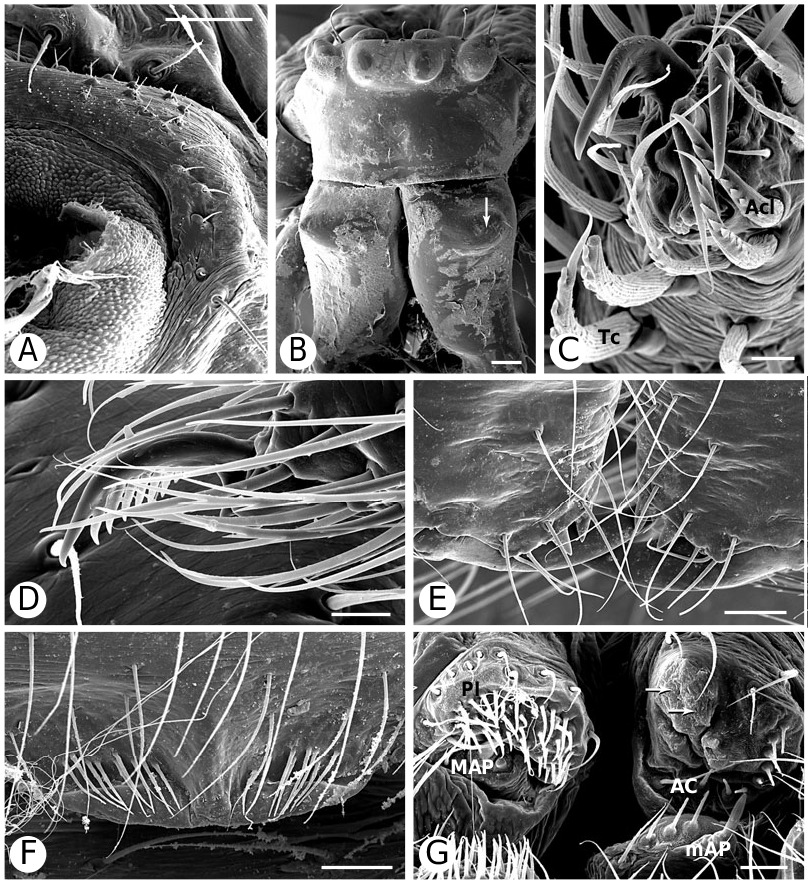
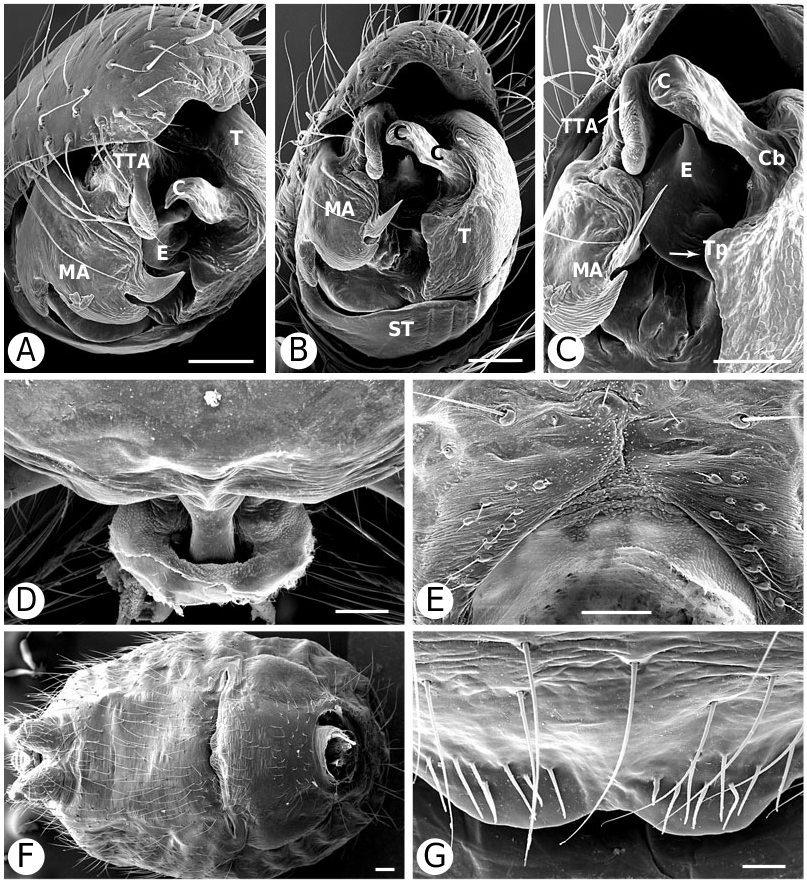
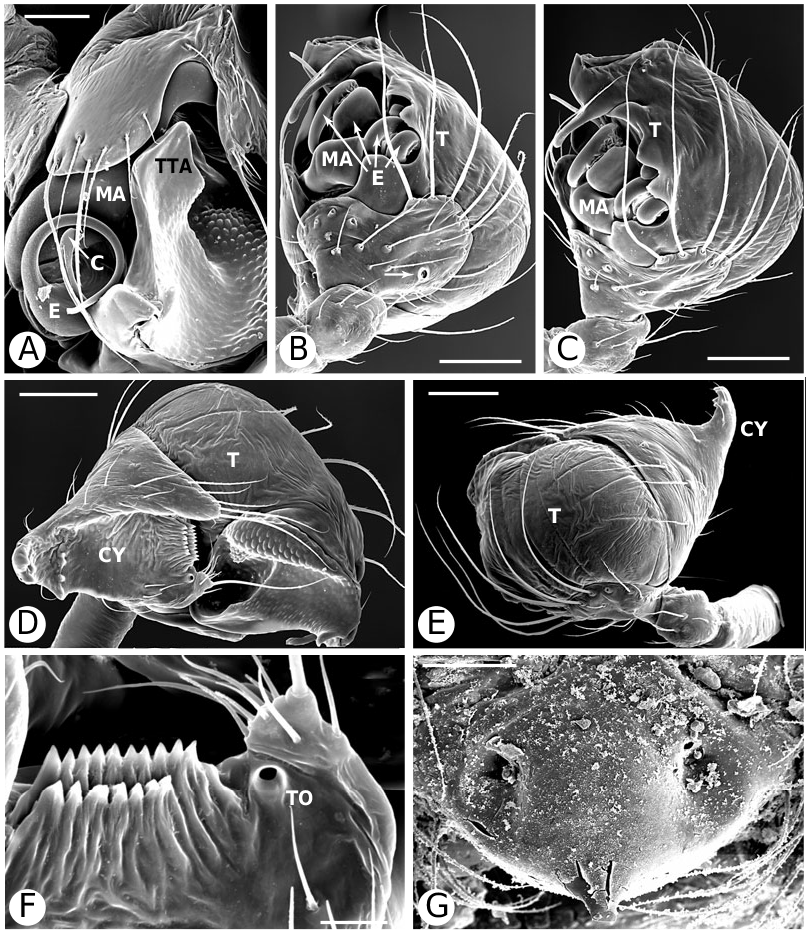
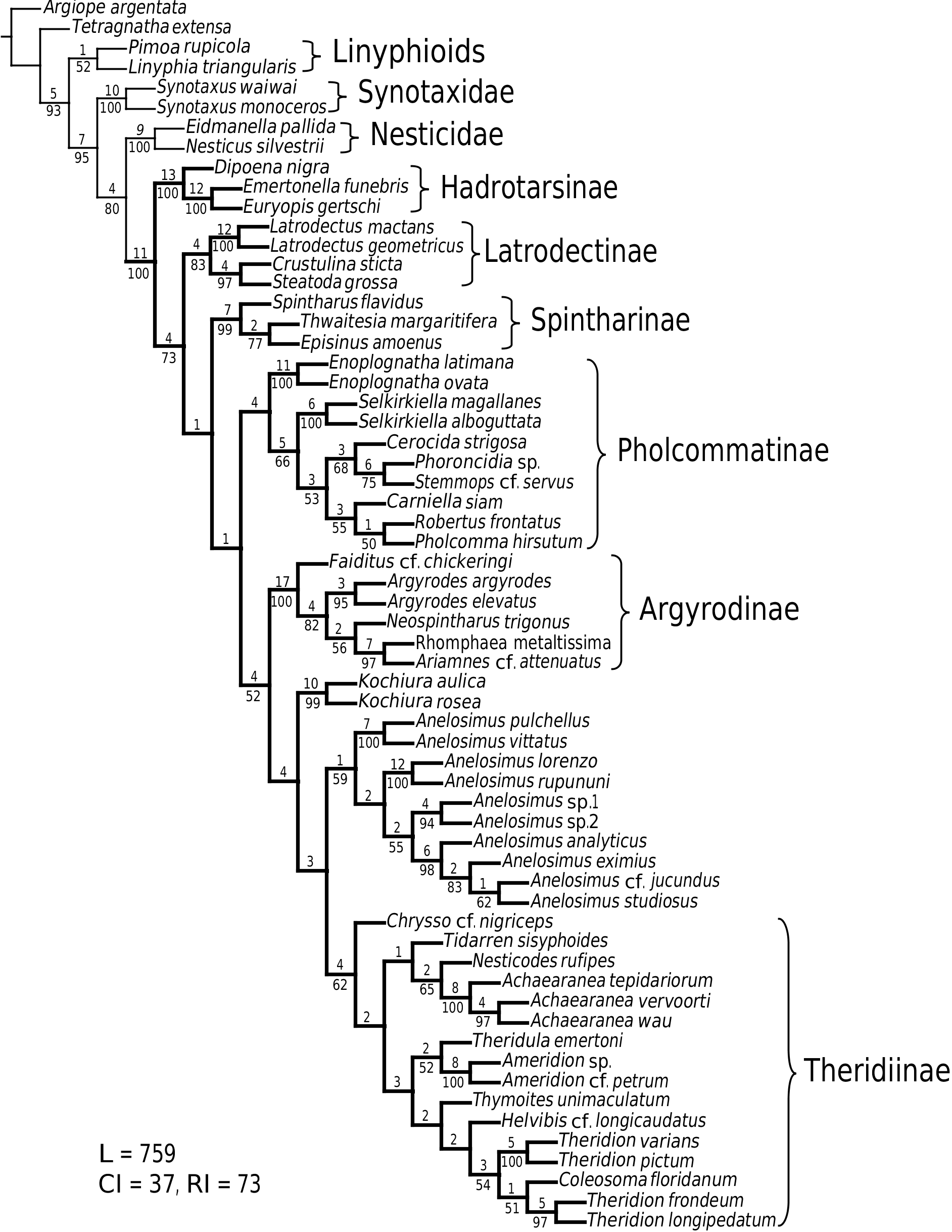
As predicted by Forster et al. (1990), the results strongly refute the monophyly of Anelosimus sensu lato (which included Kochiura and Selkirkiella ) ( Fig. 102 View Figure 102 ). None of the 17 Chilean species currently placed in Anelosimus seem to belong there; they differ in many key characteristics from the type species A. eximius , such as plesiomorphic retention of the hooked cymbial notch ( Fig. 67B, D, E View Figure 67 ), hooded MA ( Fig. 67D View Figure 67 ), and colulus ( Fig. 68F View Figure 68 ). Some Chilean ‘ Anelosimus ’ are clearly pholcommatines ( A. alboguttatus and A. magallanes are here transferred to the resurrected pholcommatine Selkirkiella , clade 39), based for example, on cymbial hook position and its structural details (34, 35, 37, Fig. 67D View Figure 67 ), embolus position (89, Fig. 67B View Figure 67 ), and spinneret spigot morphology (216, Fig. 68D, E View Figure 68 ) (see Taxonomy section). Others belong to a clade containing the European A. aulicus , whose placement in Anelosimus is also refuted ( A. aulicus and A. roseus are sister to the lost colulus clade and are here transferred to the resurrected Kochiura , clade 26; see Taxonomy section).
As circumscribed here, the monophyly of Anelosimus rests on only one unambiguous synapomorphy: ridges on the surface of the epigynal plate (3, Figs 19B View Figure 19 , 21G View Figure 21 , 93F View Figure 93 ). Bremer support for the clade is low (1), the clade is sensitive to data perturbation, and making clade 23 sister to Theridiinae involves only one step. On the other hand, clade 18, which contains the ‘classical’ new world Anelosimus , is well supported (Fig. 104). Many new species have come to light (for example, from Madagascar; pers. observ.), and the taxonomy will probably change as new data are added.
Theridiinae is well defined (see below) and easily diagnosed, including all nonhadrotarsine theridiids without any trace of colulus. The inclusion of Anelosimus View in CoL in Theridiinae would muddle the diagnosis of the subfamily and add to what is already the largest theridiid subfamily. As the position of Anelosimus View in CoL outside the lost colular setae clade is corroborated by molecular evidence ( Arnedo et al., 2004), I do not include it in Theridiinae . The placement of Kochiura View in CoL is uncertain; here it is outside the lost colulus clade, while in Arnedo et al. (2004) it is sister to Anelosimus View in CoL . The subfamily placement of Kochiura View in CoL and Anelosimus View in CoL thus awaits future combined analysis. The results suggest dual origin of quasisociality within Anelosimus View in CoL (see ‘Sociality’, below).
THERIDIINAE (LOST COLULAR SETAE CLADE): CLADE 15
Yoshida (2001a: 158) defined Theridiinae based on two synapomorphies: ‘paracymbium of male palpus hooded’ and ‘colulus usually absent’ and synonymized Spintharinae with it. Yoshida’s Theridiinae is apparently not monophyletic and is redefined here, while Spintharinae is again recognized (see above).
Theridiinae (including at least Achaearanea , Ameridion , Chrysso , Helvibis , Nesticodes , Theridion , Theridula, Thymoite and Tidarren ) is well supported in this study. It is unambiguously defined by: a reduction in both retrolateral (18, with some reversals within the subfamily) and prolateral trichobothria (19) on the male palpal tibia ( Figs 85B View Figure 85 , 86B View Figure 86 ), epiandrous gland spigots spread over genital plate (169, Figs 58E View Figure 58 , 76F View Figure 76 , 77G View Figure 77 ), and absence of colular setae (no trace of colulus, 174, Fig. 78A View Figure 78 ). In addition, most species in this clade have notably long and thin legs.
Other nonhadrotarsine theridiid genera lacking the colulus probably belong here, including Achaearyopa Barrion & Litsinger, 1995 , Cabello Levi, 1964 , Cephalobares O. P.-Cambridge, 1870, Cyllognatha L. Koch, 1872 , Echinotheridion Levi, 1963 , Exalbidion Wunderlich, 1995 , Histagonia Simon, 1895 , Jamatid- ion Wunderlich, 1995, Keijia , Macaridion Wunderlich, 1992 , Molione Thorell, 1892 , Neottiura , Nipponidion Yoshida, 2001 , Paratheridula Levi, 1957 , Propostira Simon, 1894 , Rugathodes Archer, 1950 , Sardinidion Wunderlich, 1995 , Simitidion Wunderlich, 1992 , Takayus Yoshida, 2001 , Tekellina Levi, 1957 , Wamba O. P.-Cambridge, 1896, and perhaps others.
This group is similar to the colulus-less group discussed by Levi & Levi (1962). About half of theridiid species belong to Theridiinae ; the group is therefore relatively poorly understood. Most genera still have no identified synapomorphies and relatively few appear to be monophyletic. The name Theridiinae refers to Simon’s tribal group Theridieae, which was based on Theridion and related genera.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Anelosimus
| Agnarsson, Ingi 2004 |
Anelosimus
| Simon 1891 |
Anelosimus
| Simon 1891 |
Anelosimus
| Simon 1891 |
Anelosimus
| Simon 1891 |
Anelosimus
| Simon 1891 |