
Cosmolaelaps lignicolus
|
publication ID |
https://doi.org/10.11646/zootaxa.4232.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:6F270C13-A16A-45CD-AB95-545C619E5FED |
|
DOI |
https://doi.org/10.5281/zenodo.5698403 |
|
persistent identifier |
https://treatment.plazi.org/id/8309879F-F93A-2035-B9AE-F8CDFCB8FD36 |
|
treatment provided by |
Plazi |
|
scientific name |
Cosmolaelaps lignicolus |
| status |
|
Cosmolaelaps lignicolus (G. & R. Canestrini, 1882) comb. nov.
( Figs 1–31 View FIGURES 1 – 2 View FIGURES 3 – 7 View FIGURE 8 View FIGURES 9 – 12 View FIGURE 13 – 15 View FIGURES 16 – 21 View FIGURES 22 – 24 View FIGURES 25 – 26 View FIGURES 27 – 28 View FIGURES 29 – 34 )
Laelaps lignicola G. & R. Canestrini, 1882a: 74. Laelaps lignicola G. & R. Canestrini, 1882b: 72.
Hypoaspis lignicola .— Canestrini, 1885: 89; Oudemans, 1902: 24; Oudemans, 1903: 129; Moreira, 2014: 409. Laelaps ( Iphis) lignicola .—Berlese, 1892: 70, 2.
Laelaps lignicola .— Tipton, 1960: 299.
Gymnolaelaps lignicola .— Bernini et al., 1995: 27.
Material examined. The material of C. lignicolus studied here was collected mostly by the senior author exclusively from nests of Lasius emarginatus (Olivier) , and where it is not indicated, this material is deposited in the Collection of Lentate sul Seveso Civic Museum, Monza & Brianza, Italy.
Specimens examined: 2 females, Veneto, Padova, rotting wood, other collecting data unknown, Berlese Acaroteca ( Centro di ricerca per l'agrobiologia e la pedologia, Florence ), slide 44/38 [original label: COLL. BERLESE— Hypoaspis ♀ —( Gymnolaelaps )— lignicola —cotipi ( G. R. Can.)— Padova legno guasto] ; 2 females, Liguria, Genova , date unknown, A. Dodero coll., Berlese Acaroteca, slide 50/34 [original label: COLL. BERLESE— Gymnolaelaps ♀♀ — lignicola —( G. R. Can.)—Genova—( Dodero!)] ; 2 females, Liguria, Genova , date unknown, A. Dodero coll., Berlese Acaroteca, slide 50/35 [original label: COLL. BERLESE— Gymnolaelaps — lignicola ♀♀ —( G. R. Can.)—Genova—( Dodero!)] ; 1 female, Liguria, Genova , date unknown, A. Dodero coll., Berlese Acaroteca, slide 208/50 [original label: COLL. BERLESE— Hypoaspis ♀ — ( Gymnolaelaps )— lignicola —rotta ( G. R. Can.)—Genova—( Dodero!)] ; 1 female, Liguria, Genova , date unknown, A. Dodero coll., Berlese Acaroteca, slide 209/2 [original label: COLL. BERLESE— Hypoaspis ♀ — ( Gymnolaelaps )— lignicola —( G. R. Can.)—Genova—( Dodero!)] ; 10 females, 1 male and 1 deutonymph, Lombardia, Milano province, Solaro , 4 October 2009 ; 16 females and 1 male, Puglia, Foggia province, Vieste, Foresta Umbra , 20 August 2013 ; 85 females and 4 males, Lombardia, Como province, Albavilla, Alpe del Vicerè , 10 November 2013 ( 10 females and 1 male deposited in the Acarological Collection, Department of Plant Protection, Yazd Branch, Islamic Azad University, Iran; 4 females deposited in the Arthropods Collection of Natural History Museum, Athens, Georgia, United States) ; 6 females, Lombardia, Como province, Eupilio, Monte Cornizzolo , 9 March 2014 ; 5 females, Lombardia, Milano province, Senago , 23 March 2014 ; 3 females, Lombardia, Milano province, Bollate, Castellazzo , 30 March 2014 ; 6 females, 1 male and 2 deutonymphs, ITALY, Lombardia, Como province, Limido Comasco , 25 May 2014 ; 39 females, 36 males and 14 deutonymphs, Emilia Romagna, Ferrara province, Comacchio, Lido di Pomposa , 1 June 2014 ( 8 females and 4 males deposited in the Acarological Collection, Department of Plant Protection, Yazd Branch, Islamic Azad University, Iran; 2 females and 1 male deposited in Canestrini collection at Zoological Museum, University of Padua, Italy; 2 males deposited in the Arthropods Collection of Natural History Museum, Athens, Georgia, United States) ; 13 females, 1 male and 3 deutonymphs, Toscana, Livorno province , San Vincenzo, S. Carlo, 21 July 2014 ; 4 females and 1 male, Piemonte, Alessandria province, Castelnuovo Scrivia , 3 August 2014 ; 9 females, Lombardia, Varese province, Lonate Pozzolo , 30 March 2014, M. Galuppi coll. ; 4 females, Toscana, Livorno province, Isola d'Elba, Monte Perone , 27 July 2015 ; 3 females and 1 male, Piemonte, Alessandria province, Mongiardino Ligure, Salada , 20 March 2016 .
Diagnosis (adults and deutonymph). Idiosoma prolonged posteriorly, with only tarsal segment of the fourth legs protruding beyond its posterior margin; all idiosomatic setae simple, without a discernible asymmetric swelling. Dorsal shield highly convex in adults, with some setae in marginal and submarginal position ( r2–r5, s1– s3, s6, S1–S5 and extra pairs x1–x3) barely discernible in dorsal view (even in partially pressed specimens); most dorsal shield setae relatively short, slightly decreasing in length from anterior to posterior positions in opisthonotal region; opisthonotum slightly hypertrichous in deutonymphs; setae Z5 thickened, blade-like, barbed in distal end; lyrifissures idJ2 and idJ4 not discernible in adults; three ( x1–x3) and one ( x1) extra pairs of marginal setae respectively in adults and deutonymph. Opisthogastric setae short. Sternal shield distinctly wider than long in female, with irregular lobes arising anteriorly to st 4 in deutonymph. Genital shield distinctly enlarged behind coxae IV. In the male: setae Px generally absent; outer lobes of internal malae without fimbriae; seta av on telefemur II inserted on a well-developed apophysis, seta av on genu and tibia II, and setae mv, pv2 and av2 on tarsus II, thickened, spine-like, and shorter than in female.
Description (adults and deutonymph).
Female. Figs 1–12 View FIGURES 1 – 2 View FIGURES 3 – 7 View FIGURE 8 View FIGURES 9 – 12 , 16–18 View FIGURES 16 – 21 , 22, 24 View FIGURES 22 – 24 , 29–30 View FIGURES 29 – 34 ; 10 specimens measured. Idiosoma oval-shaped, prolonged posteriorly, with only tarsal segment of the fourth legs protruding beyond its posterior margin; 617 ± 13 (600–640) long and 474 ± 8 (460–490) wide, ratio length/width 1.30 ± 0.03 (1.24–1.35).
Dorsal idiosoma ( Figs 1–7 View FIGURES 1 – 2 View FIGURES 3 – 7 , 29–30 View FIGURES 29 – 34 ). Dorsal shield entire, completely covering dorsal surface, highly convex, with some setae in marginal and submarginal position ( r2–r5, s1–s3, s6, S1–S5 and extra pairs x1–x3) barely discernible in dorsal view (even in pressed specimens) ( Fig. 1 View FIGURES 1 – 2 ); with 43 pairs of setae, 23 pairs on podonotum ( j1– j6, z1–z6, s1–s6, r2–r5; r6 ventrally placed) including a marginal supernumerary pair ( x1) posterior to setae s6, and 20 pairs on opisthonotum ( J1–J5, Z1–Z5, S1–S5) including three pairs of extra setae ( Px1–Px3) ( Zx1–Zx3 of other authors) between J and Z series and two marginal supernumerary pairs, at level of setae S1 ( x2) and between setae S1 and S2 ( x3); setae J2–J4 very unstable in position (in some cases, one or both setae of these pairs apparently absent); with a variable number (4–7) of unpaired setae ( Jx) always present between J series (in some specimens, setae Jx between J4 and J5 appearing as a supplementary pair); all setae simple, without a discernible asymmetric swelling, setiform and smooth ( Figs. 29–30 View FIGURES 29 – 34 ), except j1, z1 and Z5, thickened, j1 and z1 acuminate, Z5 ( Fig. 2 View FIGURES 1 – 2 ) blade-like and barbed in distal end (in some specimens setae J4, J5, Z4 and Jx, between J4 and J5, slightly barbed); most setae not long enough to reach base of next posterior seta, j1 and z1 the shortest, other podonotal setae subequal in length, opisthonotal setae slightly decreasing in length from anterior to posterior positions, with Z5 the shortest: j 1 18–19, j2 34–38, j5 39–45, z 1 14–16, z2 38–43, z3 35–41, z4 44–49, z5 43–46, s5 46 –51, x1 36 –40, x 3 30–34, J1 38–43, J2 39–41, J4 34–38, Jx 33–35, J 5 31–39, Z1 43–46, Z2 38–41, Z3 34–38, Z4 39–41, Z 5 20–23, Px1 39–46, Px2 36–46, Px3 34–40; with 23 pairs of discernible pore-like structures, 12 on podonotum ( idj1, idj3, idz3, idr5, idj6, ids6, idx1, gdj2, gdj4, gdr4, gdz5, gdz6; gds4 not discernible) and 11 on opisthonotum ( idZ1, idS1, idS2, idJ1, idPx, idS3, idZ4, idJ5, idZ5, gdx2, gdS3; gdZ3 not discernible), with idJ2 and idJ4 apparently absent; dorsal surface smooth, ornamented throughout with irregular polygonal pattern, less regular in the central region of shield.
Ventral idiosoma ( Figs 8–12 View FIGURE 8 View FIGURES 9 – 12 ). Tritosternum with subrectangular base well separated from anterior margin of sternal shield, base longer 39 ± 3 (35–44) than basal width 20 ± 1 (16–21), and two free pilose laciniae. Presternal area with faint lines of sclerotisation, without platelets well-developed. Sternal shield well sclerotised, wider than long, 67 ± 3 (63–70) long, 126 ± 3 (123–131) wide; with anterior margin well defined, medially almost straight, and antero-lateral corners pointed, concave on posterior margin; with three pairs of smooth setae ( st1 40–45, st 2 45–50, st 3 40–43) and two pairs of slit-like lyrifissures, adjacent to st1 and between st 2 and st 3 setae; surface delicately ornamented with irregular polygonal pattern. Endopodals II–III fused with sternal shield, endopodal platelets III–IV distinct and well-developed, boomerang-shaped, not fused anteriorly to sternal shield, with deeper adaxial portion, posteriorly connected with sclerites of the sperm access system. Metasternal platelets absent; setae st 4 (30–33) situated on soft cuticle covering adaxial portion of the endopodal platelets III–IV, poroids iv3 on soft integument, well separated from endopodal platelets. Genital shield flask-shaped, enlarged behind coxae IV, 317 ± 7 (305–323) long, 201 ± 3 (198–205) wide; widely separated from anal shield and extending posteriorly to level of Jv1 setae; with rounded posterior margin and hyaline anterior margin, anteriorly reaching beyond posterior margin of sternal shield; bearing only setae st5 (34–35); setae Zv1 (15–16) and Jv1 (18–21) close to the edge of the shield, on unsclerotised cuticle; genital lyrifissures iv5 on soft integument beside genital setae; surface of the shield ornamented with irregular polygonal pattern, basally composed of large cells, the outer edges of the upper cells forming an inverted u- or v-shaped line. Peritrematal shields only with antiaxial and post-stigmatic sections developed, not extending behind coxa IV, free from exopodals and fused to dorsal shield only in a very small section anterior to z1; peritreme extending anteriorly beyond coxae I, up to level between s1 and z1; each shield bearing five discernible pore-like structures (a lyrifissure ip and a gland pore gp at level of coxa II, two lyrifissures ip and a gland pore gp on post-stigmatic section); a small pore-like structure within peritreme at level of coxa I. Setae r6 (12–15) on unsclerotised integument between peritrematal shields and antero-lateral margins of dorsal shield, about at level of stigmata. Exopodal platelets I–II absent, exopodals II–III present, although weakly sclerotised, generally posteriorly not connected with well-developed exopodal platelets III–IV, the latter ones extending along posterior margin of coxae IV, but not expanded posteriorly in podal elements; glands gv2 present on posterior rim of exopodals III–IV. Anal shield subtriangular, about as long as wide, 106 ± 5 (100–113) long, 105 ± 4 (100–110) wide; anal opening situated about at mid-level of shield; para-anal setae (23–25) longer than unpaired post-anal seta (20–21); pores gv3 on lateral margins of the anal shield, situated from the mid-level of anal opening to just behind its posterior margin; cribrum extending anteriorly along lateral margins of shield beyond insertion of post-anal seta; with some sculptural lines on anterior half. Opisthogastric unsclerotised integument with 15 pairs of setae, excluding a variable number of setae UR (chaetotaxy with some variation; Figs 9–12 View FIGURES 9 – 12 ), and five pairs of lyrifissures ivp (excluding genital pores iv5); Jv1 and Jv2 pairs with setae widely separated from each other, displaced posteriorly ( Jv1 about at level of Zv2); all setae smooth and needle-like, subequal in length (except Jv2, the longest), not reaching base of next posterior seta ( Jv 1 18 – 21, Jv 2 23–25, Jv 3 18–21, Jv 4 13–15, Jv 5 13–15, Zv 1 15– 16, Zv 2 15–18, Zv 3 11–15, Zv 4 11–13, Zv 5 10–13, R 1 11–15, R 2 10–13, R 3 9–13, R 4 10–13, R 5 10–13, UR 9–13, except seta UR between R1 and post-stigmatic section of peritrematal shield, 15–19, when present always longer than setae R and other setae UR); metapodal platelets elongate, oriented obliquely, 42 ± 5 (35–49) long; one pair of small parapodal elements variable in shape; two pairs of rod-shaped platelets close to Zv1, one larger, between Zv1 and lateral margin of the genital shield (generally not fused with shield), the other one tiny, external to Zv1.
Gnathosoma ( Figs 16–18 View FIGURES 16 – 21 ). Epistome triangular, with anterior margin mostly smooth, with sparse isolated denticles ( Fig. 16 View FIGURES 16 – 21 ). Hypostome with three pairs of smooth and needle-like setae, h1 (43–46) shorter than h3, h2 (21–26) the shortest, h3 (60–64) the longest; palpcoxal setae (33–38) smooth and needle-like; deutosternal groove usually with six-seven rows of denticles, each bearing 4-10 small teeth; corniculi horn-shaped, parallel, reaching about middle of palp-femur and about four times as long as basal width ( Fig. 18 View FIGURES 16 – 21 ). Internal malae bilobed, inner lobe narrow, pointed and serrated, outer lobe wide, bearing well-developed fimbriae with rounded tips ( Fig. 18 View FIGURES 16 – 21 ). Labrum blade-like, pilose, considerably longer than corniculi and internal malae. Palp chaetotaxy normal for the genus; apotele 2-tined, tines of unequal length; all setae smooth and needle-like, except setae al on femur and al1 on genu, both stout and spur-like; al2 about twice length of al1 ( Fig. 18 View FIGURES 16 – 21 ). Chelate-dentate chelicerae welldeveloped, movable digit bidentate, fixed digit multidentate generally with four robust teeth approximately equally-sized (some of which can be partially or totally fused together) and three small subapical teeth; dorsal seta short, arthrodial membrane extended into a brush-like structure; inner face of chelicera with an irregular window of weakly sclerotised integument, connected with a narrow scissure arising dorsally from base of fixed digit ( Fig. 17 View FIGURES 16 – 21 ).
Legs ( Figs 22, 24 View FIGURES 22 – 24 ). Leg II stouter than other legs ( Fig. 24 View FIGURES 22 – 24 ). Chaetotaxy normal for free-living Laelapidae and genus Cosmolaelaps ; leg I: coxa 0 0/1 0/1 0, trochanter 1 0/2 1/1 1, femur 2 3/1 2/3 2, genu 2 3/2 3/1 2, tibia 2 3/2 3/1 2; leg II ( Fig. 24 View FIGURES 22 – 24 ): coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/2 1, genu 2 3/1 2/1 2, tibia 2 2/1 2/1 2; leg III: coxa 0 0/1 0/1 0, trochanter 1 0/2 0/1 1, femur 1 2/1 1/0 1, genu 2 2/1 2/1 1, tibia 2 1/1 2/1 1; leg IV ( Fig. 22 View FIGURES 22 – 24 ): coxa 0 0/1 0/0 0, trochanter 1 0/2 0/1 1, femur 1 2/0 1/1 1 ( pd about half as long as length of ad1 and ad2), genu 2 2/1 3/0 1, tibia 2 1/1 3/1 2 ( av and pv thicker and longer than other setae); tarsi II–IV with 18 setae, 3 3/2 3/ 2 3 + setae mv and md; all setae simple and needle-like; all pre-tarsi with a well-developed ambulacral stalk, a pair of claws and three rounded pulvillar lobes, extensively projecting beyond claws.
Spermathecal apparatus ( Fig. 8 View FIGURE 8 ). Sperm access system composed of a complex of sclerites, situated at level of coxa IV and continuous with endopodals III–IV; proximal section of tubulus sclerotised and enlarged; other structures unsclerotised.
Male ( Figs 13–15 View FIGURE 13 – 15 , 19–21 View FIGURES 16 – 21 , 23 View FIGURES 22 – 24 , 31 View FIGURES 29 – 34 ; 5 specimens measured). Idiosoma slightly more oval-elongate than in female; 575 ± 33 (530–615) long and 415 ± 19 (395–435) wide, ratio length/width 1.39 ± 0.10 (1.28–1.51). Dorsal idiosoma ( Figs 13–14 View FIGURE 13 – 15 ). Dorsal shield narrower than in female, with setae and pore-like structures of some series noticeably closer than in female (i.e. idPx2, Z4 and idZ4); extension, shape, chaetotaxy and pore-like structures of dorsal shield as in female, except for setae Px1–Px3 generally absent.
Ventral idiosoma ( Figs 14–15 View FIGURE 13 – 15 ). Tritosternum and presternal area as in female. Sterno-genital, ventro-anal and endopodal shields fused in a well-developed holoventral shield, with anterior margin well defined, prominent at level of genital opening, and posterior portion of variable extension and shape; shield 475 ± 24 (455–510) long and 254 ± 16 (230–275) wide (behind coxae IV), bearing 10 pairs of simple and needle-like setae, excluding circumanal setae ( st1–st5, Zv1–Zv2, Jv1–Jv3), 5 pairs of lyrifissures ( iv1 slit-like; a pair of lyrifissures ivp about at level of Jv1) and glands gv3, situated about at mid-level of anal opening; setae Zv2 and Jv3 on or well removed from lateral margins of shield or on unsclerotised integument (depending on the shape and lateral extension of shield at level of these setae); Jv1 and Jv2 more anteriorly and medially located than in female; st1–st3 and st5 subequal in length, st4 shorter than other sternal setae ( Table 1 View TABLE 1 ); relative length of setae Zv1–Zv2 and Jv1–Jv3 highly variable ( Zv1 with greater variation; Table 1 View TABLE 1 ); para-anal setae longer than unpaired post-anal seta (as in female); cribrum as in female; shield throughout with polygonal ornamentation, except posteriorly to anal opening. Peritrematal shields only with antiaxial and post-stigmatic sections developed, not extending behind coxa IV and free from exopodals (as in female), broadly fused to dorsal shield in their anterior portion, about to level of r2 ( Fig. 14 View FIGURE 13 – 15 ); peritreme extending anteriorly beyond coxae I, up to level of s1 ( Fig. 14 View FIGURE 13 – 15 ); peritrematal and post-stigmatic pore-like structures as in female. Setae r6 as in female. Exopodal shields as in female; exopodals III–IV connected with holoventral shield. Opisthogastric unsclerotised integument with 10 pairs of setae, excluding a variable number of setae UR, and 5 pairs of lyrifissures ivp; all setae smooth and needle-like, subequal in length ( Zv3– Zv 5 11–16, Jv4–Jv 5 11– 18; seta UR between R1 and post-stigmatic section of peritrematal shield, when present always longer than setae R and other setae UR, as in female).
Gnathosoma ( Figs 19–21 View FIGURES 16 – 21 ). Epistome ( Fig. 19 View FIGURES 16 – 21 ) and hypostome as in female. Inner lobes of internal malae as in female, outer lobes narrow, serrated, without fimbriae ( Fig. 21 View FIGURES 16 – 21 ). Labrum as in female. Palp chaetotaxy as in female. Movable digit of chelicera with one large tooth, spermatodactyl digitiform, longer than movable digit and with rounded tip; fixed digit multidentate, with two-three distal teeth (subapical tooth clearly larger) and a well separated and oblique more proximal masticatory ridge, composed by three-four small teeth; inner face of chelicera with a narrow scissure arising dorsally from base of fixed digit (without the window of weakly sclerotised integument present in female); other cheliceral structures as in female ( Fig. 20 View FIGURES 16 – 21 ).
Legs ( Figs 23 View FIGURES 22 – 24 , 31 View FIGURES 29 – 34 ). Chaetotaxy as in female. Leg II stouter than in female; seta av on femur II inserted on a well-developed tubercle and shorter than in female; seta av on genu and tibia II, and setae mv, pv2 and av2 on tarsus II, thickened, spine-like, and shorter than in female ( Figs 23 View FIGURES 22 – 24 , 31 View FIGURES 29 – 34 ).
Deutonymph. Figs 25–28 View FIGURES 25 – 26 View FIGURES 27 – 28 , 32–33 View FIGURES 29 – 34 ; 6 specimens measured. Exoskeleton soft, depigmented. Idiosoma narrower and with posterior end more acute than in adults, 539 ± 19 (520–570) long and 356 ± 21 (330–390) wide, ratio length/width 1.52 ± 0.04 (1.46–1.58).
Dorsal idiosoma ( Figs 25–26 View FIGURES 25 – 26 , 28 View FIGURES 27 – 28 , 32–33 View FIGURES 29 – 34 ). Dorsal shield entire, posteriorly not completely covering dorsal surface, significantly less convex and laterally less expanded than in adults, with a narrow incision along each lateral margin, just anteriorly to S1 and extending medially about up to level of z6; with 40 pairs of setae, 23 pairs on podonotum ( j1–j6, z1–z6, s1–s6, r2-r5; r6 ventrally placed, as in adults) including a marginal supernumerary pair ( x1) posterior to s6 setae, and 17 pairs on opisthonotum ( J1–J5, Z1–Z5, S1–S5) including two pairs of extra setae ( Px2–Px3) between J and Z series (setae x2, x3 and Px1 absent); with a variable number (10–16) of unpaired setae ( Jx) always present between J series; disorganized additional setae x (0–3) may be present between setae Px and J series; in some cases setae r on unsclerotised integument (i.e. r3, Fig. 25 View FIGURES 25 – 26 ; r4, Fig. 28 View FIGURES 27 – 28 ); S3 on the lateral margin of shield or on unsclerotised integument ( Fig. 25 View FIGURES 25 – 26 ); Z3 close to lateral margin of shield, but well removed from it; shape of setae as in adults ( Figs 26 View FIGURES 25 – 26 , 32–33 View FIGURES 29 – 34 ); some setae relatively longer than in adults and reaching base of next posterior seta (i.e. z3 and s5 reaching beyond insertion of z4 and z6, respectively), j1 and z1 the shortest podonotal setae (as in female), opisthonotal setae slightly decreasing in length from anterior to posterior positions with Z5 the shortest (as in female): j 1 16–18, j3 38–40, j4 35–44, j5 38–43, z 1 13–15, z2 43–48, z4 43–50, z5 38– 43, s4 51 –58, J1 36–40, J 2 30–35, J 3 30–34, J 4 25–30, Jx (about at level of idJ1) 28–31, Jx (about at level of Px3) 23–28, Jx (between J4 and J5) 18–23, J 5 20–29, Z1 38–45, Z2 35–43, Z4 33–40, Z 5 19–21, Px 2 25–33, Px 3 25–30, x 30–33; with at least 21 pairs of pore-like structures, 10 on podonotum ( idj1, idj3, idj6, idz3, idr5, ids6, gdj2, gdj4, gdz5, gdr4; gdz6 and idx1 apparently absent) and 11 on opisthonotum ( idZ1, idJ1, idS1, idPx, idJ2, idS2, idS3, idJ4, idZ4, idJ5, idZ5; gdx2 and gdS3 apparently absent); dorsal surface ornamented throughout with irregular polygonal pattern, less marked in the central region of shield.
Ventral idiosoma ( Figs 27–28 View FIGURES 27 – 28 ). Tritosternum and presternal area as in adults. Sternal shield weakly sclerotised, with anterior margin and antero-lateral corners not well defined (continuous with presternal lines of sclerotisation), and posterior margin broadly rounded; longer than wide, 220 ± 8 (208–230) long (anterior measuring point at level of first pair of lyrifissures), 100 ± 6 (93–105) wide (widest point just anterior to insertion of st3); shield than narrows sharply behind two pronunced and irregular lobes, arising anteriorly to st4; with at least three pairs of setae simple and smooth, subequal in length ( st 1 31–38, st2 33–38, st 3 30–38), and three pairs of lyrifissures (the first and second pairs oval, respectively behind st1 and between st 2 and st 3, the third pair the smallest, circular, posterior to st3 and on lateral margins of shield); st4 shorter than other sternal setae (18–21), generally on the tip of sternal lobes or out of shield, on soft integument ( Fig. 27 View FIGURES 27 – 28 ); ornamentation of shield not well discernible. Distinct endopodal platelets absent. Peritrematal shields absent, only with two small weakly sclerotised portions, the proximal one bearing a gland pore gp; peritreme extending anteriorly beyond coxae I, up to level between s1 and z1 (as in female), and posteriorly not behind coxa IV; a gland pore gp posteriorly to stigma, on unsclerotised integument (other post-stigmatic pore-like structures not discernible). Setae r6 as in adults. Exopodal platelets absent; glands gv2 not discernible. Anal shield subtriangular, longer than wide, 93 ± 2 (89–96) long, 77 ± 4 (73–83) wide (at level of pores gv3); anal opening situated about at mid-level of shield; para-anal setae longer than unpaired post-anal seta (as in adults); pores gv3 not well discernible (often circumscribed by integument sclerotised in a spiral fashion), on lateral margins of anal shield, at level of anal opening as in adults; cribrum and sculptural lines as in female. Opisthogastric unsclerotised cuticle with 16 setae (including st5), in addition to a variable number of setae UR, and 5 pairs of discernible circular lyrifissures ivp; position of Jv1 and Jv2 setae as in male; all setae smooth and needle-like, subequal in length ( st 5 19–23, Jv 1 14–18, Jv 2 18–20, Jv 3 18–19, Jv 4 14–16, Jv 5 13–15, Zv 1 13–15, Zv 2 13–16, Zv 3 13–16, Zv 4 13–14, Zv 5 11–14, R1–R 5 10–16, UR 11–14); metapodal platelets present, proportionally smaller than in female, elongate and oriented at 45°; parapodal elements or other platelets absent.
Gnathosoma . All gnathosomal structures as in female, except for palpal trochanter and femur, distinctly shorter than in female; epistome generally not discernible; all morphological details of the cheliceral dentition exactly as in female.
Legs. Leg II stouter than others (as in adults). Chaetotaxy as in female.
Remarks. In adults of C. lignicolus the dorsal shield is highly convex and laterally expanded, often extending onto the ventral surface in mounted specimens. This prevents the observation of marginal and submarginal morphological details, which in this species include some setae and pore-like structures very close to lateral margins of shield ( Fig. 3 View FIGURES 3 – 7 ). In deutonymphs, the dorsal shield is significantly less convex and expanded than in adults (as shown by the position of the setae r and S, on the lateral margin of the shield or on unsclerotised cuticle) and very marginal structures of the adults are absent (with the exception of the setae x1; Figs 25 View FIGURES 25 – 26 , 28 View FIGURES 27 – 28 ). These observations suggest that the origin of these structures might be correlated with the lateral expansion of the shield in adults.
Based on our observations mostly in adult females, and in contrast to what has been reported in literature for other laelapid mites, in C. lignicolus the correct chaetotaxy for femur I seems be 2 3/1 2/3 2 (three setae ad and two setae pd). In fact, in this species the most proximal dorsal seta of femur I is inserted more or less in the middle of the segment, but in most specimens it appears shifted more toward the anterior (2 3/1 2/3 2) than the posterior margin (2 2/1 3/3 2) of the segment. Likewise, in C. lignicolus the more proximal ventral seta of trochanter II (1 0/ 1 0/2 1, one seta av and two setae pv) and the only ventral seta of femur IV (1 2/0 1/1 1, one seta pd; Fig. 22 View FIGURES 22 – 24 ) appear closer to the posterior than the anterior margin of the relative leg segment.
Males and females of C. lignicolus show differences at species-level for some secondary sexual characters. Among others (i.e. different shape and chaetotaxy of leg II), of special interest are some features of internal malae and opisthonotal setation. Based on descriptions from literature and direct examination of some European species [i.e. representatives of the C. cuneifer -group and C. tuberculatus ( Mašán, 1992) ], in Cosmolaelaps species whose females display external lobes of the internal malae with well-developed fimbriae, these structures are generally equally developed also in males. Interestingly, the females of C. lignicolus have external lobes with fimbriae welldeveloped, whereas the males of the same species have external lobes merely fringed, with true fimbriation absent ( Figs 18, 21 View FIGURES 16 – 21 ). We have also observed fimbriae well-developed and fimbriae absent, respectively in female and male specimens of the C. vacuus -group collected from Italy [see also figures 200-HW and 200-HM in Hirschmann et al. (1969) for C. serratosimilis (Bernhard) , a junior synonym of C. vacuus , syn. by Bregetova (1977)]. The existence of this secondary sexual dimorphism might be due to specific differences in the biology of the sexes but, in the absence of further information, it is difficult to propose a hypothesis. In regard to the chaetotaxy of the opisthonotum, in the females setae J2–J4 seem be unstable in position and presence ( Figs 1 View FIGURES 1 – 2 , 4–7 View FIGURES 3 – 7 ), whereas in the males, a slightly narrower idiosoma and a reduction of the distance between the setae and pore-like structures of some series (i.e. clearly discernible in the case of idPx2, Z4 and idZ4) are associated with the apparent absence of the setae Px1–Px3 ( Fig. 13 View FIGURE 13 – 15 ).
Finally, it is interesting to note that in deutonymphal stage described here, the fimbriation of internal malae is developed as in females, and the opisthonotum is slightly hypertrichous (with a greater number of setae Jx than in adults and the presence of supernumerary setae x). In one deutonymph has been observed a dorsal seta with a well distinct bulging base ( Fig. 33 View FIGURES 29 – 34 ). However, it is not possible conclude whether this seta should be considered an aberrant seta or a rare attribute of the species.
Ecological notes and distribution. The previous records of Cosmolaelaps lignicolus documented its presence in three localities of peninsular Italy, where this species was respectively collected from a soaked tree (G. & R. Canestrini 1882), under stones, from decomposing organic substrate ( Canestrini 1885), and from unspecified habitat (this latter record is reported in Castagnoli & Pegazzano 1985 and it is relative to some specimens collected by the Italian entomologist Agostino Dodero near the city of Genoa).
All the new specimens of C. lignicolus have been exclusively collected from anthills of Lasius emarginatus . This ant is a very thermophilous Euro-Caucasian, Mediterranean and sub-Mediterranean species, inhabiting lowland and submontane areas. Its nests can be found in warm and rocky natural habitats, as well as in rural and urban disturbed areas ( Seifert 2007). The senior author has observed Cosmolaelaps lignicolus in anthills under stones or most often within logs and stumps, mainly in various forest habitats, where the most abundant samples were also obtained. The species was also collected from nests in open areas, albeit with series of fewer specimens. The relationship between this laelapid mite and its ant host is demonstrated by further observations: (1), C. lignicolus has never been found by the senior author in many samples obtained from rotting logs or other substrates not colonised by ant species; (2), most specimens have been observed in deeper sections of the anthills, where the cocoons of the ant host were present; in cases where the nests were not dissected in depth, individuals of C. lignicolus have been observed to come out occasionally and slowly from the crevices of inner and more humid parts of the anthill; (3), deutonymphs have also been found in anthills, suggesting that a large part of the life cycle is accomplished in this habitat; (4), C. lignicolus has not been found in anthills of some syntopic species of L. emarginatus and with very similar ecology [i.e. L. lasioides (Emery) , L. niger (L.) and L. platythorax Seifert ] or in those of other formicid species, suggesting a high host specificity.


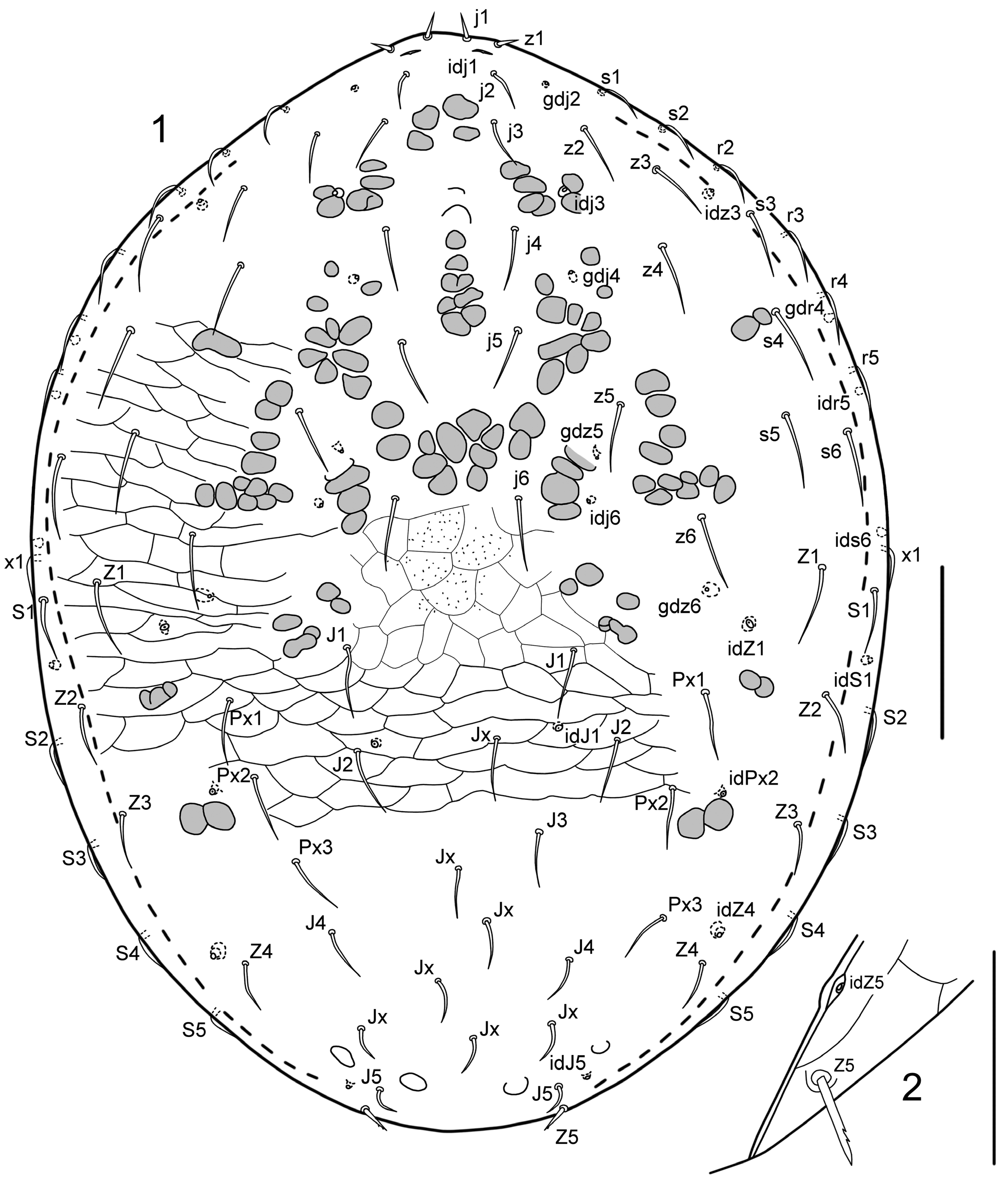
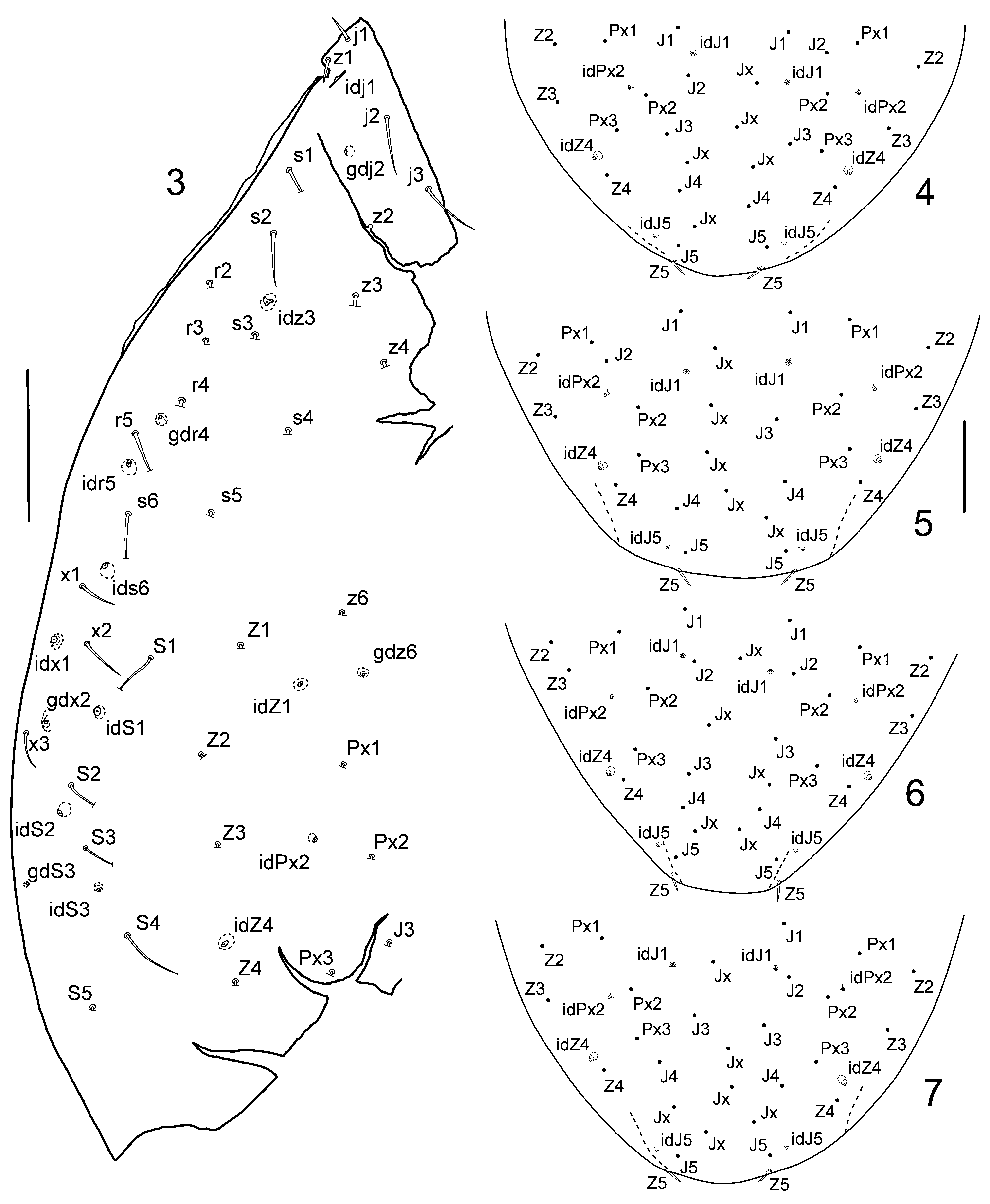

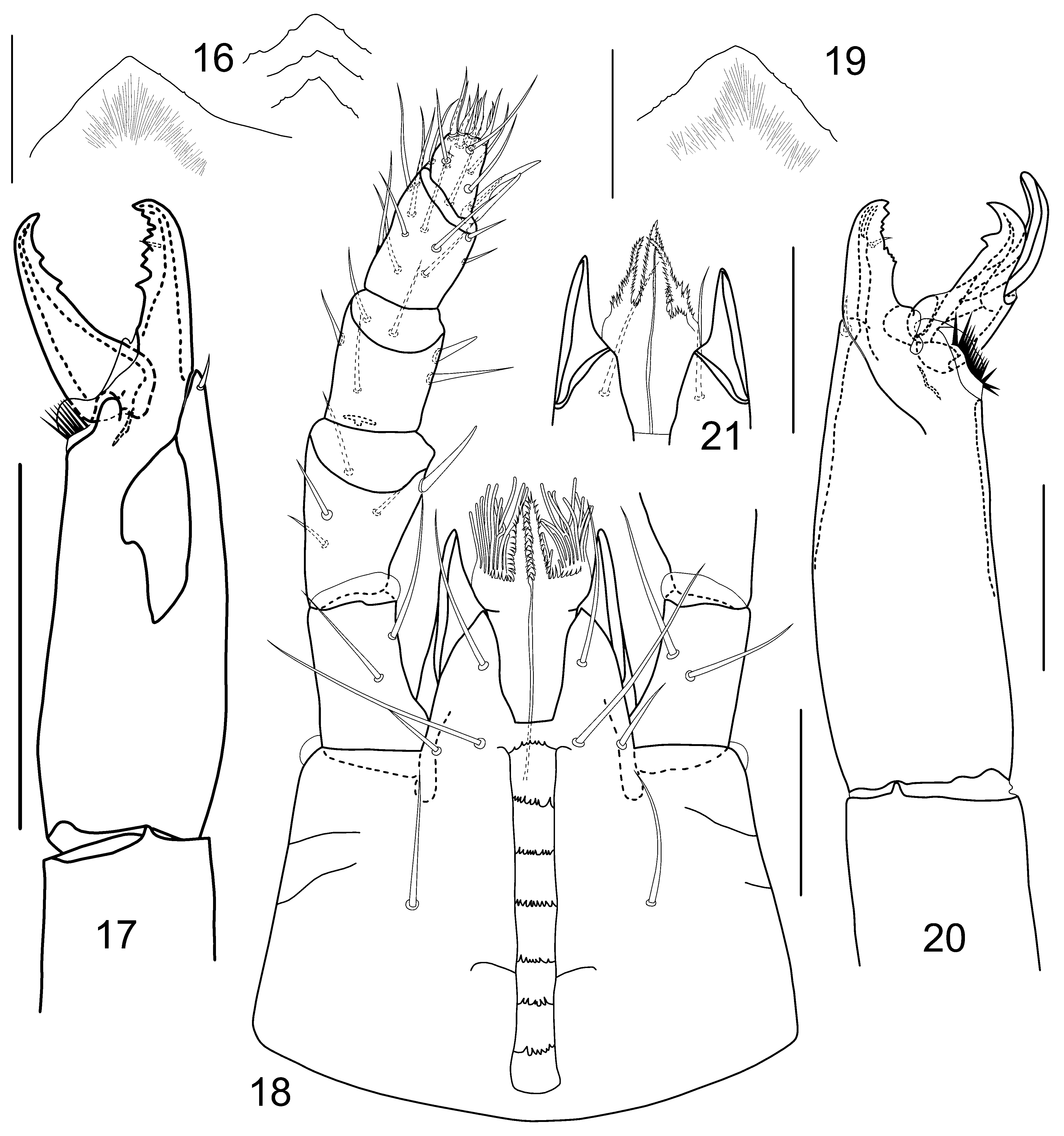


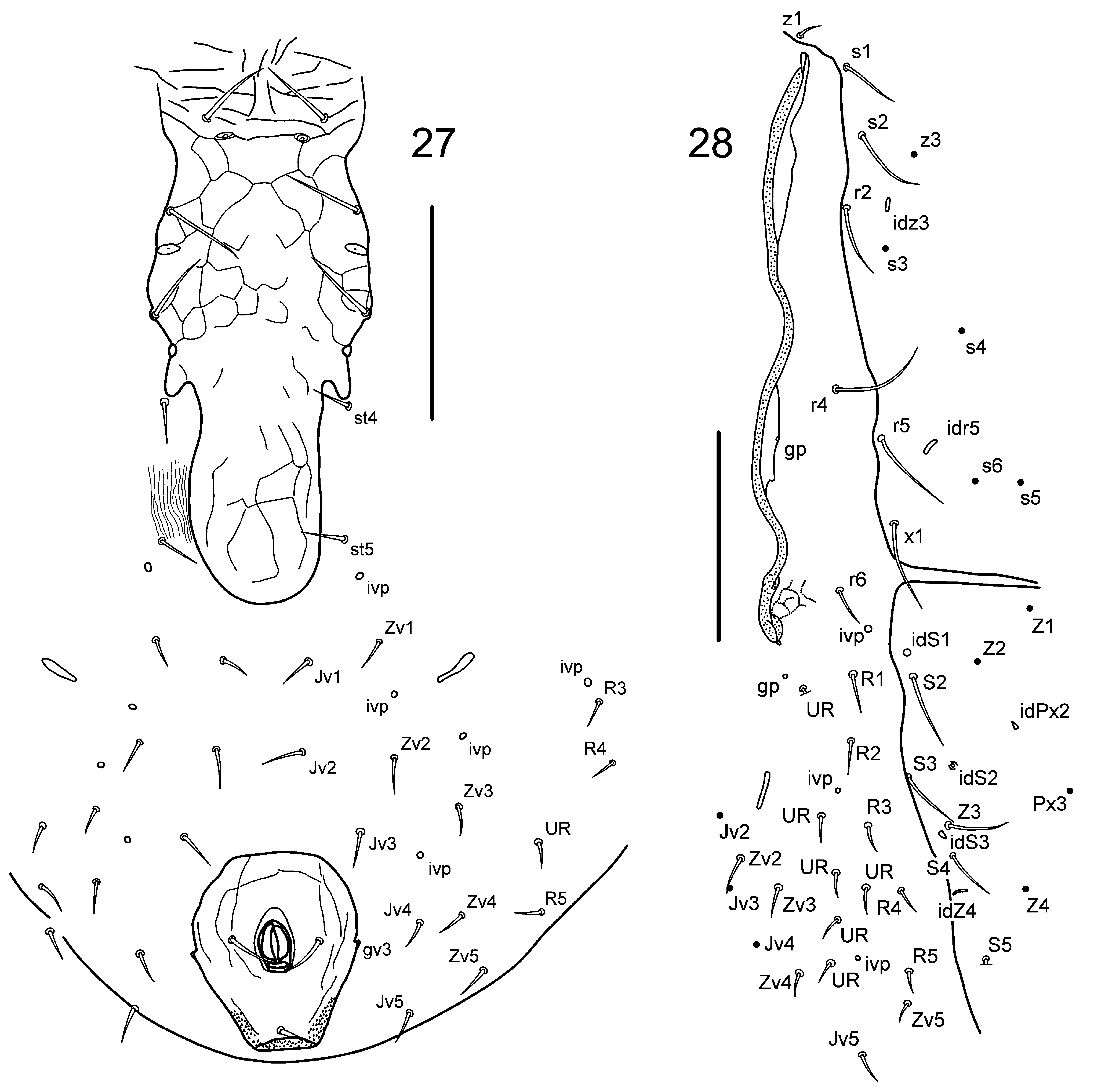
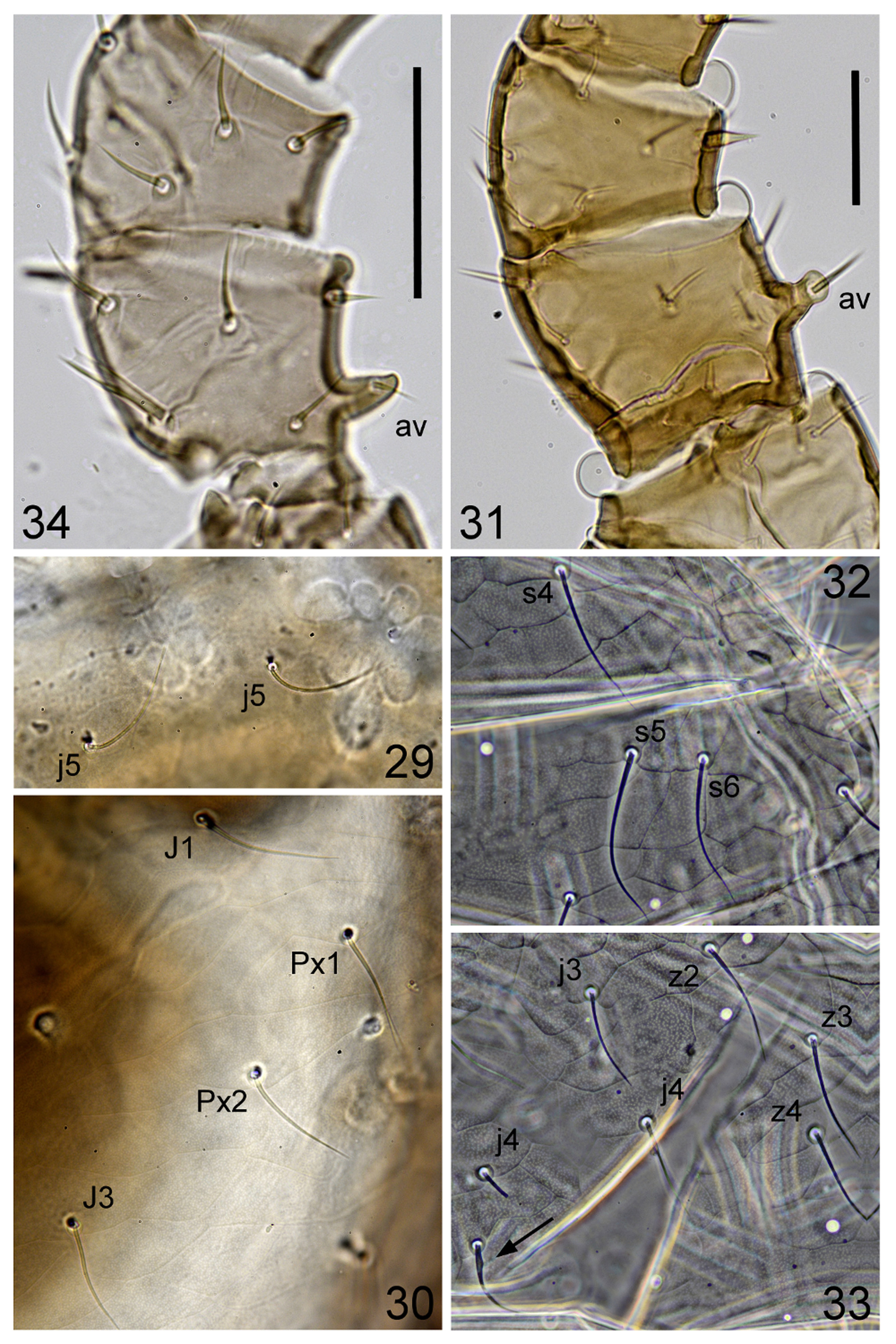
TABLE 1. Length of holoventral shield setae in five males of Cosmolaelaps lignicolus.
| st1 | st2 | st3 | st4 | st5 | Zv1 | Zv2 | Jv1 | Jv2 | Jv3 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 28 | 28 | 28 | 23 | - | 30 | 39 | 36 | 39 | 29 |
| 2 | 2 8 | 2 6 | 2 8 | 2 3 | 2 5 | 1 6 | 4 4 | 4 0 | 4 5 | 3 6 |
| 3 | 2 5 | 2 8 | 2 6 | 2 0 | 2 5 | 2 3 | 5 1 | 4 5 | 5 0 | 3 8 |
| 4 | - | 26 | 26 | 21 | 24 | 21 | 31 | 30 | 34 | 33 |
| 5 | 2 5 | 2 6 | 2 6 | 2 1 | 2 5 | 2 9 | 4 5 | 4 5 | 5 1 | 3 9 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cosmolaelaps lignicolus
| Plumari, Massimo & Joharchi, Omid 2017 |
Gymnolaelaps lignicola
| Bernini 1995: 27 |
Laelaps lignicola
| Tipton 1960: 299 |
Hypoaspis lignicola
| Oudemans 1903: 129 |
| Oudemans 1902: 24 |
| Canestrini 1885: 89 |
Laelaps lignicola
| Canestrini 1882: 74 |
| Canestrini 1882: 72 |