
Geocenamus conicaudatus, Ghaderi, Reza & Karegar, Akbar, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4079.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:87A3A1AB-48DC-4E3D-9948-0EA039660D46 |
|
DOI |
https://doi.org/10.5281/zenodo.6060896 |
|
persistent identifier |
https://treatment.plazi.org/id/830E87A8-FFF0-FFD8-EB9B-9CB3F3E1BA2A |
|
treatment provided by |
Plazi |
|
scientific name |
Geocenamus conicaudatus |
| status |
|
Description of Geocenamus conicaudatus n. sp.
( Fig. 1 View FIGURE 1 , 5 View FIGURE 5 A–E, 6 A–E, 8 F)
= Scutylenchus sp. A in Ghaderi et al. (2014) = Merlinius rugosus apud Hashim, 1980
Measurements. See Table 1 View TABLE 1 .
Female. Body straight to open C-shaped. Cuticle prominently annulated; annuli about 1.6–2.3 µm wide at midbody. Lateral fields 6–10 µm wide or 27–35% of body width; with six incisures along most of the body length, but only four post-phasmid; outer or inner bands occasionally irregularly areolated. Longitudinal striae 32–34 at midbody (two specimens). Distance from stylet knob base to lip constriction 10–13 cuticular annuli. Head hemispherical, set-off from the rest of the body by a distinct constriction, 4–5 µm high and 7–9 µm wide at base; perioral disc distinct in some individuals. Head with five or six annuli. Cephalic framework slightly sclerotized, basal margins not refractive. Anterior and posterior cephalids indistinct. Stylet slender, 1.9–2.5 times as the lip region width; conus 8–11 µm, as long as shaft ( m = 47.6–54.4); knobs 3–4 µm across, rounded, laterally directed; rarly anterior surfaces slightly sloping backward. Orifice of dorsal pharyngeal gland 1.5–2.5 µm from the base of stylet. Deirids not observed. Hemizonid two to three annuli long, one to six annuli anterior to the secretoryexcretory pore. Median pharyngeal bulb oval, with prominent valve plates; distance from the anterior end to its center 53–67 µm. Isthmus slender, 1.0–1.4 times as the length of the basal bulb. Nerve ring 73–107 µm from the anterior end. Basal pharyngeal bulb cylindrical, length/width ratio 1.6–3.1, offset from intestine; dorsal gland nocleus at middle of basal bulb and subventral glands nuclei slightly posterior. cardia prominent, rounded.
Vulva transverse slit, sunken or flush with the body surface, with inner double epiptygmata. Vagina 22–39 % body-width long, moderately sclerotized, usually dilated inwards as its walls curve out to uterus. Spermathecae rounded, with rounded sperm cells. Ovaries outstretched, with a single row of oocytes, except in multiplication region. Rectum about half of the anal body width, no post-anal intestinal sac present. Tail conical, tapering gradually, ending to a pointed to finely rounded non-annulated terminus. Tail terminal hyaline region three to five µm thick. Sub-cuticular layer refractive, but with variable thickness from about one to four µm. Phasmids 11–26 µm posterior to the level of anus, in anterior half of the tail.
Juveniles similar to females in general, but with four incisures in the lateral field.
Male. Similar to female in general dimensions. Stylet length and development comparable with that of females. Spicules slightly curved ventrally, with a terminal notch. Gubernaculum simple, crescent-shaped. Bursa enveloping entire tail; hypoptygma distinct. Phasmids at middle of tail length or slightly anterior.
Diagnosis and relationships. Geocenamus conicaudatus n. sp. can be characterized by having 1) hemispherical head which is set-off from body by a distinct constriction, 2) slender stylet (17–20 µm) with laterally directed knobs, 3) coarsely annulated cuticle with 32–34 longitudinal striae in addition to the six lateral field incisures, 4) conical female tail with non-annulated, pointed or finely rounded terminus, and 5) presence of males.
In having 32–34 longitudinal striae and stylet 17–20 µm long, G. conicaudatus n. sp. is comparable with G. paniculoides ( Vovlas & Esser, 1990) Brzeski, 1998 , G. tartuensis ( Krall, 1959) Brzeski, 1991 , G. siddiqii ( Mulk, 1978) Brzeski, 1991 , G. sphaerocephalus ( Ivanova, 1982) Brzeski, 1991 , G. quadrifer ( Andrássy, 1954) Brzeski , Τype pοpulatiοn (1) (2) (3) Τοtal Τοtal
Character\Pοpulatiοn
) Lebbeck tree ( Albizia lebbek ) (Ramhοrmuz-Ahvaz rοad, Khuzestan); (2) eucalyptus (Lamerd-Asalοvieh rοad, Fars); (3) tοmatο (Bοrazϳan and Abpakhsh, Bοushehr).
1991 and G. rugosus ( Siddiqi, 1963) Brzeski, 1991 . The new species differs from G. paniculoides , G. tartuensis and G. siddiqii by having non refractive basal ring-margins of the cephalic framework ( vs. refractive) and conical tail with pointed to finely rounded terminus ( vs. subcylindrical to cylindrical with rounded terminus). From G. sphaerocephalus , it can be distinguished by having a longer stylet (17–20 vs. 15–17 µm), non-areolated lateral field ( vs. areolated) and narrowly rounded (vs. broadly rounded) tail terminus. It differs from the most closely related species, G. rugosus and G. quadrifer by the tail shape (conical with finely rounded terminus vs. subcylindrical to cylindrical with broadly rounded to hemispherical terminus), demarcation of the cephalic region (set off by a constriction vs. continuous or slightly set off) and direction of the stylet knobs (laterally- vs. posteriorly- directed). It can be further separated from G. rugosus by having a shorter stylet (17–20 vs. 20–25 µm) and mode of reproduction (bisexual vs. parthenogenetic). It differs from G. ordinarius Volkova, 1993 in having slightly shorter tail (39–58 vs. 60–70 µm) and non-anulated tail terminus ( vs. annulated). Geocenamus ordinarius has weakly developed longitudinal striae, their number is unknown. The new species can be differentiated from species with similar tail morphology ( e.g., G. sobolevi ( Mukhina, 1970) Brzeski, 1991 , G. patternus Eroshenko & Volkova, 1987 , G. variabilis ( Ivanova & Shagalina, 1983) Brzeski, 1991 , G. laminatus ( Wu, 1969) Brzeski, 1991 and G. mamillatus ) by having a different number of longitudinal striae (less than 28 in the three former species and more than 50 in the two latter species).
Remarks. This species was first reported as Scutylenchus sp. A ( Ghaderi et al. 2014). Moreover, a bisexual population was reported from Jordan as Merlinius rugosus ( Hashim 1980) ; the author stated that this population differed somewhat from G. rugosus by a more prominently set-off cephalic region and a differently-shaped tail terminus, but he considered these features as intraspecific variation. We believe that the Jordanian population differs from G. rugosus , not only by the different shape of the cephalic region and tail, but also by having shorter and more slender stylet, laterally directed of the stylet knobs and the frequent presence of males. The Jordanian population can be considered as belonging to the species described in the present study, G. conicaudatus n. sp. Detailed study of the intraspecific variability in the populations of G. rugosus from Iran revealed that none of the 188 females of this species possess a conical tail with narrow terminus and/or off-set cephalic region ( Fig. 5 View FIGURE 5 & 7 View FIGURE 7 ); moreover, the female stylet was always more than 20 µm (see Table 3 View TABLE 3 & 4) and often with distinctly posteriorly directed knobs. On the other hand, males were found in low numbers in only one population of G. rugosus and with distinctly shorter stylets than the females (average 17.5 vs. 22.3 µm), but they were observed as frequently as females and with a comparable stylet length (average 18.0 vs. 18.3 µm) in all four populations of the new species described in the present study. Such differences between the stylet length of males and females (16.5 vs. 21.0 µm) can be observed in a population of G. quettensis ( Maqbool, Ghazala & Fatima, 1984) Brzeski, 1991 (now considered as a synonym of G. rugosus ) from Pakistan ( Maqbool et al. 1984).
Type habitat and Locality. The type population was collected from the rhizosphere of the Lebbek tree ( Albizia lebbeck (L.) Willd.), a shade-tree along the Ramhormuz-Ahvaz road, 30 km to Ahvaz, Khuzestan province (coordinates: 31°19'13''N, 48°40'09''E). Three other populations were collected from around the roots of eucalyptus trees (Lamerd-Asalovieh road, Fars province) and tomato plants (two populations from Borazjan and Abpakhsh counties, Boushehr province).
Type material. Holotype female, 19 paratype females, 18 males and five juveniles kept in the nematode collection of the Department of Plant Protection, College of Agriculture, Shiraz University, Iran; Three female paratypes, one male and two juveniles on slide number WT3629 and two female paratypes and one male on slide number WT 3630 in the Nematode Collection of the Plantenziektenkundige Dienst, Wageningen, The Netherlands.
Etymology. The species epithet refers to the conical tail in females.
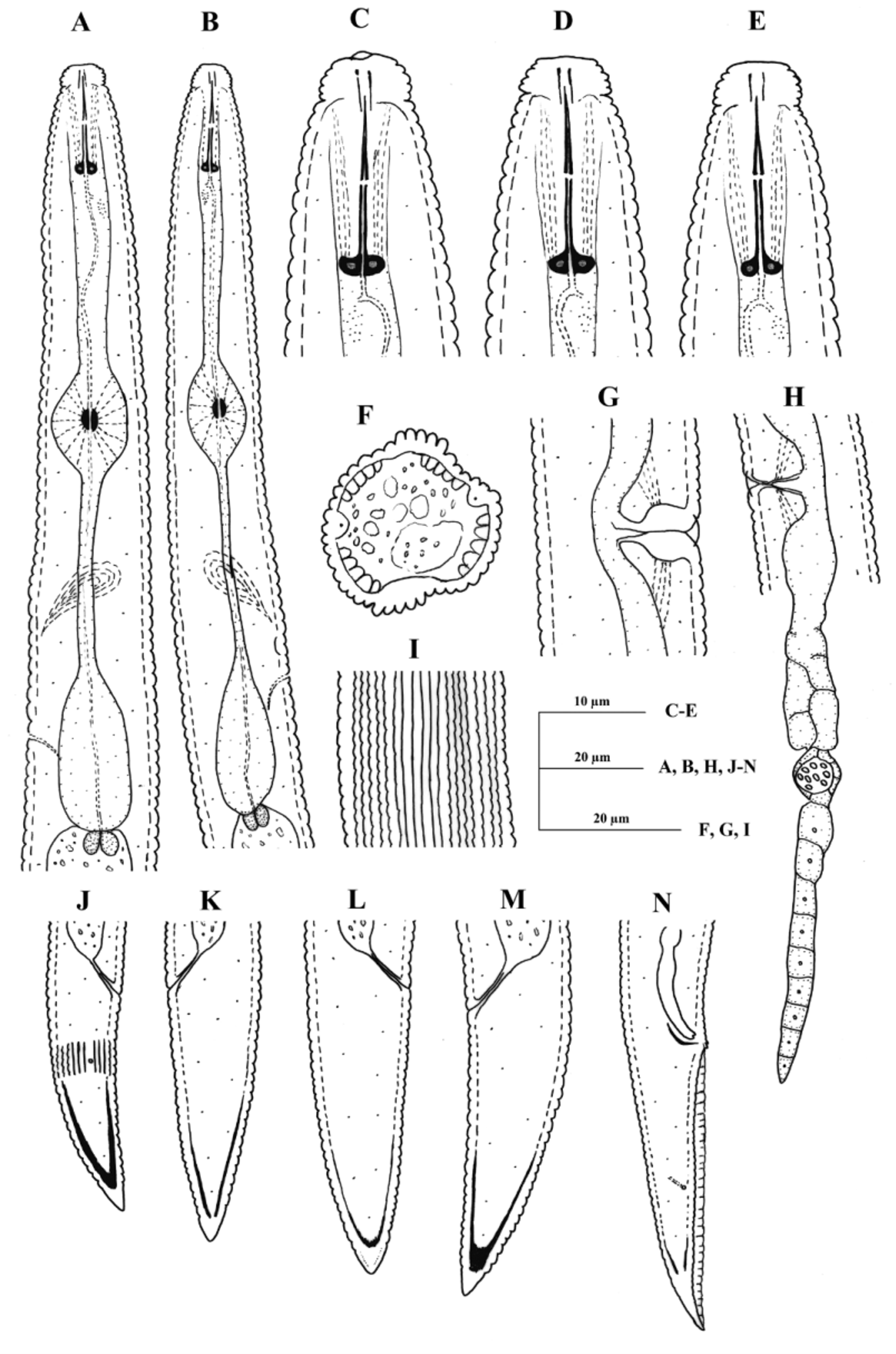
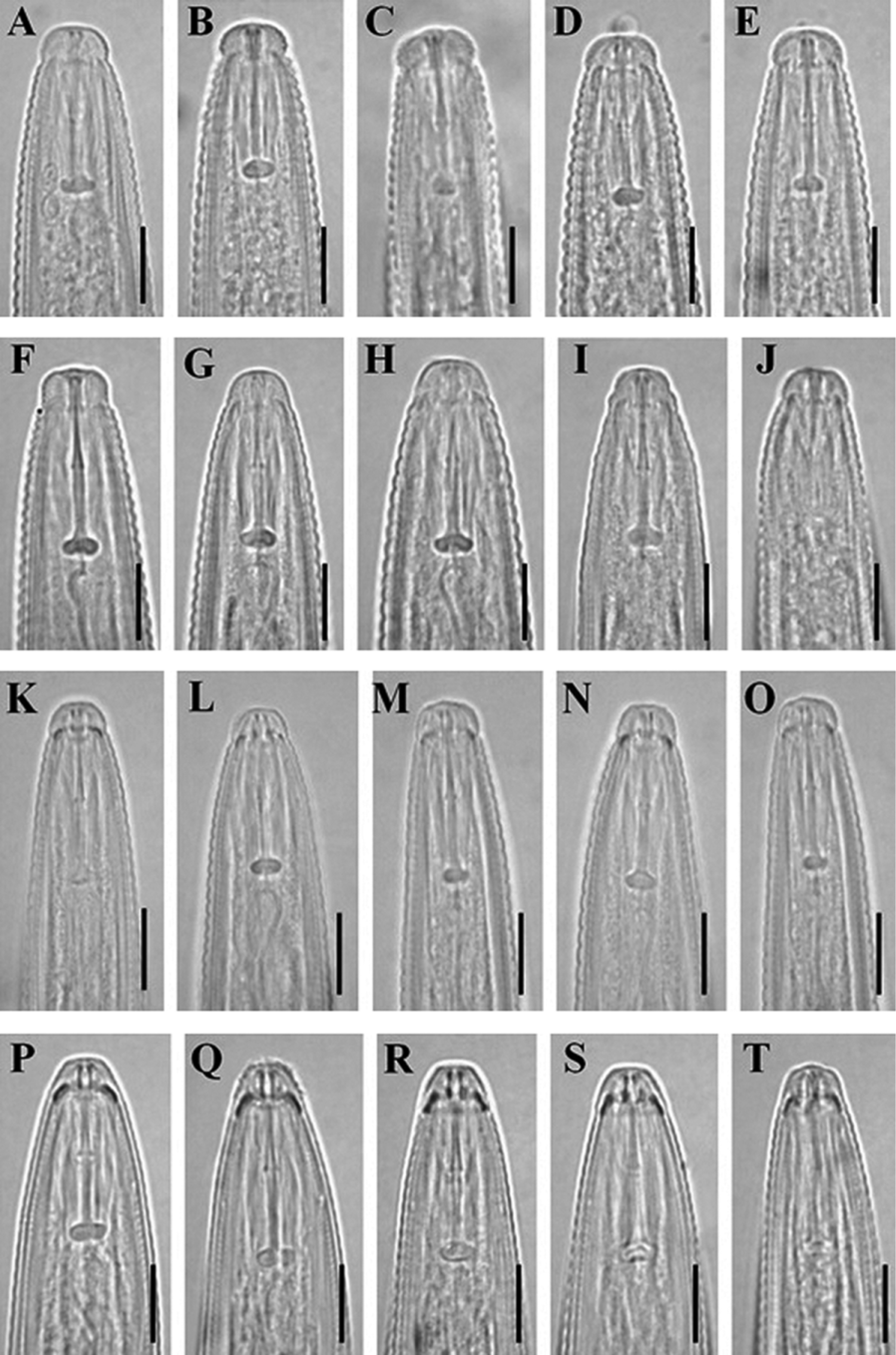
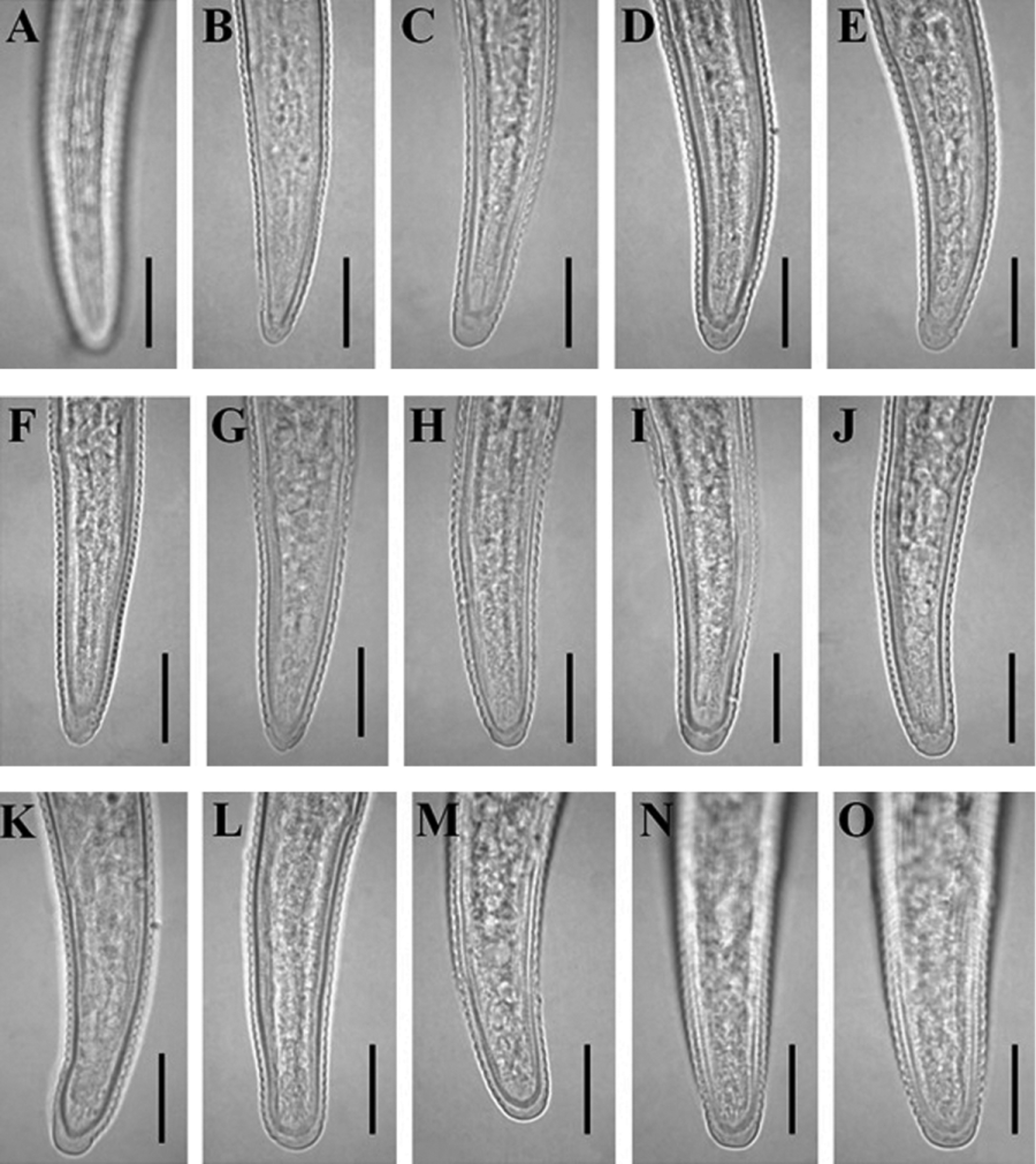
TABLE 1. Mοrphοmetric characters οf Geocenamus conicaudatus n. sp. frοm Iran (all measurements are in μm).
| n L | hοlοtype 817 | 9♀ (paratypes) 635-929 | 12♂ (paratypes) 678-920 | 12♀ 732-843 | 4♂ 781-865 | 3♀ 757-826 | 4♂ 836-903 | 25♀ 791 ± 62.7 (635-929) | Cv 7.9 | 20♂ 799 ± 70.1 (678-920) | Cv 8.8 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| a | 36.6 | 28.6-38.0 | 31.4-41.5 | 29.4-36.8 | 33.5-37.6 | 30.8-33.5 | 32.2-37.0 | 33.2 ± 2.4 (28.6-38.0) | 7.1 | 35.3 ± 2.4 (31.4-41.5) | 6.8 |
| b | 6.1 | 5.2-6.7 | 4.9-6.5 | 5.0-6.2 | 5.2-6.1 | 5.1-6.1 | 6.1-6.7 | 5.7 ± 0.4 (5.0-6.7) | 7.1 | 5.9 ± 0.5 (4.9-6.7) | 8.0 |
| c | 17.8 | 13.3-18.9 | 12.7-16.4 | 14.8-20.4 | 14.0-15.6 | 15.6-17.0 | 13.4-17.0 | 16.5 ± 1.6 (13.3-20.4) | 9.5 | 14.5 ± 1.2 (12.7-17.0) | 8.4 |
| c' | 2.7 | 2.6-3.3 | 3.0-4.2 | 2.3-3.4 | 3.4-4.0 | 2.7-2.9 | 3.1-4.2 | 2.8 ± 0.3 (2.3-3.4) | 9.1 | 3.6 ± 0.4 (3.0-4.2) | 9.8 |
| v | 56.5 | 53.2-56.8 | - | 53.1-61.2 | - | 54.2-57.7 | - | 55.8 ± 1.7 (53.1-61.2) | 3.0 | - | - |
| Stylet | 18.5 | 16.5-19.8 | 16.5-18.9 | 17.3-19.5 | 18.0-19.5 | 18.0 | 17.9-18.7 | 18.3 ± 0.8 (16.5-19.8) | 4.2 | 18.0 ± 0.8 (16.5-19.5) | 4.2 |
| m | 49.7 | 47.6-54.4 | 49.4-54.5 | 47.8-54.4 | 51.1-53.9 | 50.0-53.3 | 50.0-54.4 | 51.1 ± 2.0 (47.6-54.4) | 4.0 | 51.7 ± 1.6 (49.4-54.5) | 3.1 |
| Pharynx | 134 | 109-159 | 114-149 | 123-158 | 143-151 | 131-149 | 125-148 | 139 ± 11.3 (109-159) | 8.1 | 137 ± 10.4 (114-151) | 7.6 |
| MB | 48.1 | 45.9-49.7 | 45.6-53.3 | 46.3-52.6 | 45.6-47.8 | 44.8-47.1 | 44.9-46.9 | 47.8 ± 1.8 (44.8-52.6) | 3.7 | 47.1 ± 1.8 (44.9-53.3) | 3.8 |
| S. E. pοre | 118 | 95-138 | 101-129 | 105-123 | 106-129 | 112-124 | 108-135 | 115 ± 9.0 (95-138) | 7.8 | 116 ± 9.5 (101-135) | 8.2 |
| Basal bulb length | 26.2 | 20.4-26.2 | 23.4-26.6 | 22.0-31.2 | 23.7-31.3 | 27.4-33.6 | 22.0-30.3 | 27.5 ± 3.4 (20.4-33.6) | 12.4 | 26.4 ± 2.9 (22.0-31.3) | 11.1 |
| Basal bulb width | 12.4 | 12.4-13.7 | 10.0-13.0 | 10.0-14.3 | 10.0-13.2 | 11.6-12.6 | 9.7-12.8 | 12.1 ± 1.1 (10.0-14.3) | 8.9 | 11.6 ± 1.2 (9.7-13.2) | 10.7 |
| Head-vulva | 462 | 353-503 | - | 397-468 | - | 437-448 | - | 441 ± 34.2 (353-503) | 7.8 | - | - |
| Τail length | 46.0 | 39.3-58.0 | 48.2-59.0 | 40.4-53.2 | 54.2-58.5 | 44.5-52.8 | 53.2-65.6 | 48.2 ± 4.4 (39.3-58.0) | 9.1 | 55.3 ± 4.3 (48.2-65.6) | 7.9 |
| BW | 22.3 | 19.2-30.0 | 16.5-26.0 | 22.2-26.0 | 21.8-24.0 | 23.5-26.8 | 22.8-26.5 | 23.9 ± 2.4 (19.2-30.0) | 10.0 | 22.7 ± 2.5 (16.5-26.5) | 10.9 |
| Annuli width | 1.8 | 1.6-2.3 | 1.6-2.3 | 1.6-2.3 | 1.7-2.0 | 1.9-2.1 | 2.0-2.6 | 2.0 ± 0.2 (1.6-2.3) | 9.6 | 2.0 ± 0.3 (1.6-2.6) | 12.6 |
| Τail annuli | 24 | 20-30 | - | 17-29 | - | 24-26 | - | 24.9 ± 3.1 (17-30) | 12.5 | - | - |
| Phasmid⁄tail % | 37.6 | 28.0-48.7 | 33.5-55.5 | 24.9-48.0 | 36.9-46.7 | 29.7-39.2 | 31.6-46.1 | 35.7 ± 5.9 (24.9-48.7) | 16.5 | 42.2 ± 6.0 (31.6-55.5) | 14.3 |
| Spicules | - | - | 21.5-26.8 | - | - | - | 24.5-25.8 | - | - | 24.5 ± 1.4 (21.5-26.8) | 5.9 |
| Gubernaculum | - | - | 7.7-10.6 | - | - | - | 7.7-9.7 | - | - | 9.0 ± 1.0 (7.7-11.0) | 11.5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |