
Motmotnirmus marginellus (Nitzsch in Giebel, 1866)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4313.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A5Fdfba5-F992-44A8-84C2-1756C943C19B |
|
DOI |
https://doi.org/10.5281/zenodo.5297069 |
|
persistent identifier |
https://treatment.plazi.org/id/832187E9-FEA7-FEF3-FF74-6298FD34FE56 |
|
treatment provided by |
Plazi |
|
scientific name |
Motmotnirmus marginellus |
| status |
|
Motmotnirmus marginellus View in CoL (Nitzsch [in Giebel], 1874)
( Figs 502–508 View FIGURES 502 – 503 View FIGURES 504 – 508 )
Nirmus marginellus Nitzsch [in Giebel], 1866: 368.
Degeeriella marginella (Nitzsch [in Giebel], 1866); Harrison, 1916: 117. Brueelia marginella (Nitzsch [in Giebel], 1866); Hopkins & Clay, 1952: 58. Motmotnirmus marginellus (Nitzsch in Giebel, 1866) ; Mey & Barker 2014: 96.
Type host. Momotus momota natteri Sclater, 1858 [Ref.: Mey & Barker 2014: 96]— Amazonian motmot.
Type locality. Río Solacama, Bolivia (locality of neotype designated by Mey & Barker 2014: 97).
Other hosts. Momotus momota momota (Linnaeus, 1766) — Amazonian motmot. Momotus subrufescens Sclater, 1853 [Ref.: Carriker 1954: 200]—whooping motmot. Momotus subrufescens spatha Wetmore, 1946 [Ref.: Carriker 1954: 200]— whooping motmot. Momotus subrufescens reconditus Ridgway, 1914 [Ref.: Carriker 1954: 200]—whooping motmot. Momotus aequatorialis chlorolaemus Berlepsch & Stolzmann, 1902 [Ref.: Carriker 1954: 200]— Andean motmot.
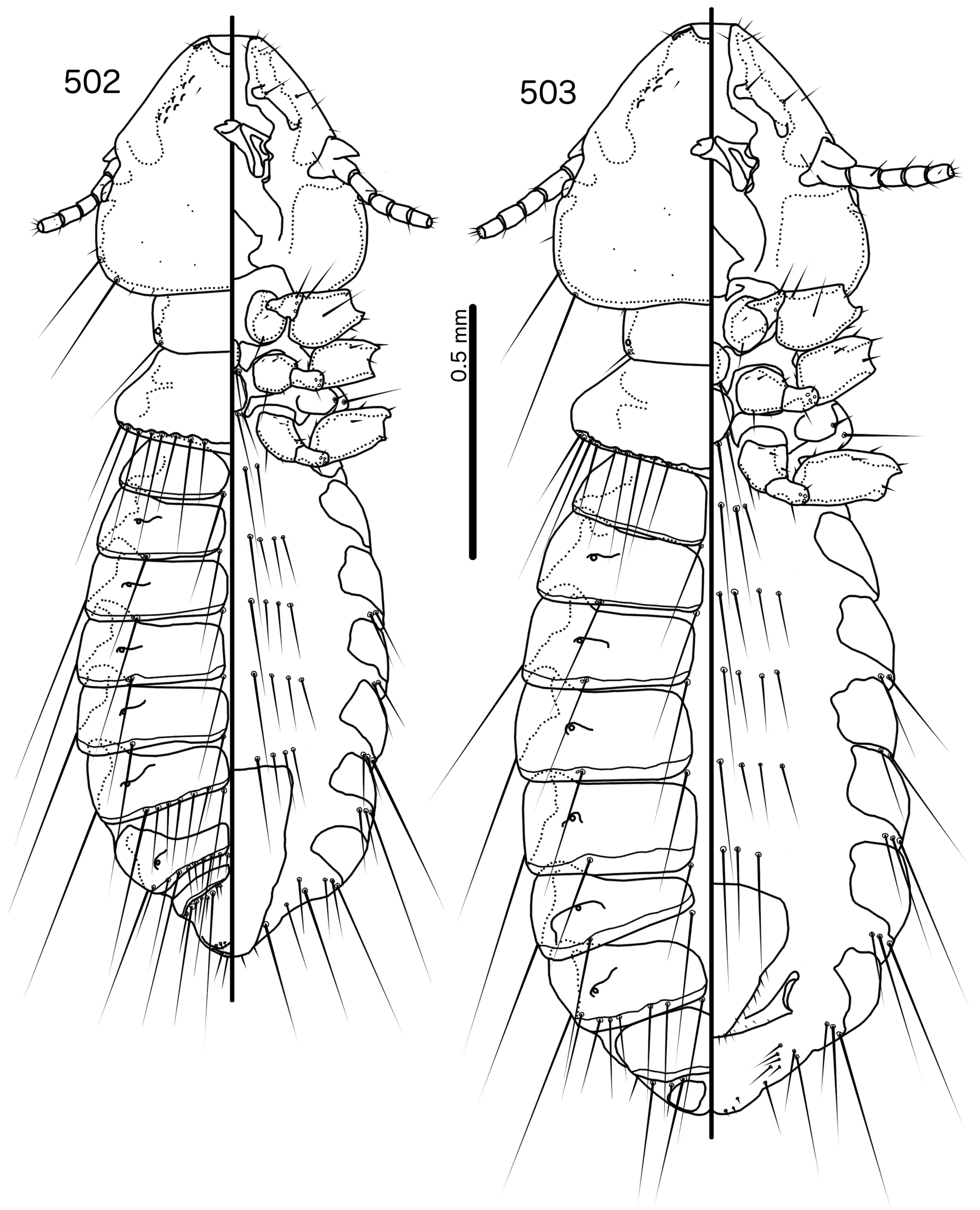
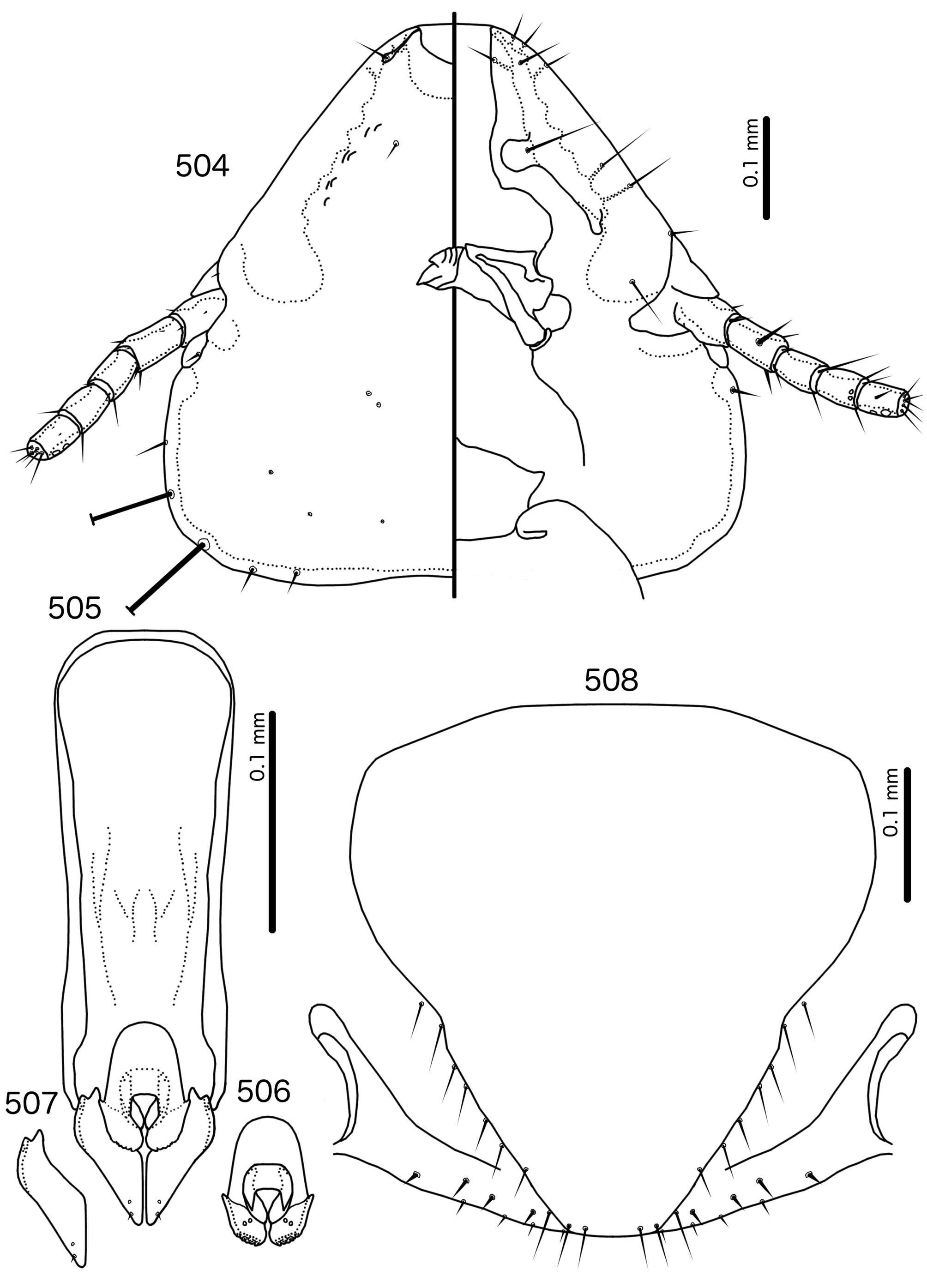
Description. Both sexes. Head shape, structure, and chaetotaxy as in genus description and Fig. 504 View FIGURES 504 – 508 . Anterior section of marginal carina of irregular width and often very hard to see; individual variation appears to be large. In some specimens displaced section of marginal carina at clypeo-labral suture cannot be seen. Preocular nodi very long, in some specimens clearly defined almost to posterior point of preantennal nodi. Postocular nodi large, rounded. Gular plate pointed medianly and laterally. Thoracic and abdominal segments as in genus description and Figs 502–503 View FIGURES 502 – 503 . Pleurites very broad.
Male. Abdominal chaetotaxy as in Table 2 and Fig. 502 View FIGURES 502 – 503 . Male genitalia as in genus description ( Figs 505–507 View FIGURES 504 – 508 ). Proximal mesosome small, rounded. Gonopore large ( Fig. 506 View FIGURES 504 – 508 ), spanning more than half of mesosome width and a third of mesosome length, narrowly open distally, with large, triangular thickenings distally. Mesosomal lobes thickened distally, rugose; 2 pmes sensilla on each lobe. Parameral heads bifid ( Fig. 507 View FIGURES 504 – 508 ). Parameral blades roughly triangular; pst1–2 as in genus description. Measurements ex Momotus momota momota (n = 6 except n = 5 for TL): TL = 1.65–1.78; HL = 0.48–0.50; HW = 0.50–0.54; PRW = 0.28–0.31; PTW = 0.45–0.49; AW = 0.59–0.65.
Female. Abdominal chaetotaxy as in Table 2 and Fig. 503 View FIGURES 502 – 503 . Subgenital plate ( Fig. 508 View FIGURES 504 – 508 ) rounded trapezoidal, reaching vulval margin but not flaring into cross-piece ( contra Mey & Barker 2014 : fig. 28). Vulval margin ( Fig. 508 View FIGURES 504 – 508 ) gently rounded with 3–4 small vms on each side, and 3–5 stout, short, but not thorn-like vss on each side; 7–10 long, slender vos; the distal 2–3 vos on posterior margin of subgenital plate, and median to vss. Measurements ex Momotus momota momota (n = 6): TL = 1.86–2.08; HL = 0.51–0.55; HW = 0.54–0.60; PRW = 0.31–0.34; PTW = 0.49–0.54; AW = 0.67–0.72.
Material examined (non-types). Ex Momotus momota momota : 2♂, 3♀, Kanaku Mountains, Rupununni, Guyana, 27 Feb. 1960, T. Clay, 173, Brit. Mus. 1961-188 ( NHML) ; 4♂, 3♀, Brazil, R. Meinertzhagen, 13454 ( NHML) ; 1♂, 1♀, Yaviza , Darien Province, Panama, 19 Nov. 1966, G.N. Lab ( OSUS) .
Remarks. Giebel (1866) compared this species with “Drosselschmaling”, meaning a slender louse on a thrush ( Turdus ). The only Nirmus from Turdus spp. he discussed were Nirmus (= Brueelia ) intermedia and Nirmus (= Guimaraesiella) marginata , and he could be referring to either of these species, most likely Gu. marginata . Giebel (1866) separated Brueelia marginellus from the slender thrush louse by its more triangular head pointed anteriorly, larger and redder preantennal nodi, and the lack of a median point of the posterior margin of the pterothorax.
Giebel (1866) did not provide any illustrations of Nirmus marginellus . Carriker (1954: figs 18–19) provided the first illustration of this species, which differs from material available to us in several respects, especially in the abdominal chaetotaxy and plates. The male genitalia in Carriker’s (1954) illustration are only barely recognizable as those of Mt. marginellus .
Carriker (1954: 200) stated that material from Momotus momota subrufescens (as Momotus subrufescens ssp.), M. momota spatha (as M. subrufescens spatha ) and M. momota chlorolaemus (as M. aequatorialis chlorolaemus ) were identical to material from M. momota momota . None of these hosts were listed by Price et al. (2003), and we tentatively recognise these species as hosts of Motmotnirmus marginellus . Carriker (1954) also noted that material from M. momota reconditus (as M. subrufescens reconditus ) was intermediate between Brueelia marginella sens. str. and Brueelia marginella xilitla , but we also tentatively recognise that bird as a host of Mt. marginellus .
Oniki & Emerson (1982: 264) considered Motmotnirmus xilitla to be synonymous with Mt. marginellus , as they could find no significant differences between them that could not “be [attributed] to age or maturity of the specimens or how they were mounted”. A small number of specimens of Mt, xilitla were available to us, including two paratypes from Momotus coeruliceps coeruliceps , which are largely indistinguishable from material from M. momota momota , except for the shape of the preantennal nodi, which narrow considerably as they approach the marginal carina. Given this difference, we tentatively retain Mt. xilitla as a separate species, until a more thorough study of motmot lice can be made. We recommend that future work on the group considers samples from a wide range of host species, populations, and geographic locations.
Mey & Barker (2014: 96) erected a neotype for Nirmus marginellus from the subspecies Momotus momota natteri , which has a very limited geographical distribution and is unlikely to be the same host subspecies as that from which Nitzsch’s original material was collected. Illustrations presented here are of material from the nominate host subspecies from Brazil. Unlike the illustrations of Mey & Barker (2014), but as in illustrations of Valim & Cicchino (2015), vsms1–2 are ventral in all specimens examined.
In the material examined by us, the abdominal chaetotaxy, dimensions, and head shape differ slightly among specimens from different host populations and localities, thus these populations may ultimately prove to be different species of Motmotnirmus . Unfortunately, the material available is too scarce to determine at present whether these different populations of Motmotnirmus on different host taxa deserve recognition as separate species, or if their differences are due to intraspecific variation.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Motmotnirmus marginellus
| Bush, Sarah E. 2017 |
Degeeriella marginella
| Mey 2014: 96 |
| Hopkins 1952: 58 |
| Harrison 1916: 117 |
Nirmus marginellus
| Giebel 1866: 368 |