
Sturnidoecus Eichler, 1944
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4313.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A5Fdfba5-F992-44A8-84C2-1756C943C19B |
|
DOI |
https://doi.org/10.5281/zenodo.5297021 |
|
persistent identifier |
https://treatment.plazi.org/id/832187E9-FF63-FF2D-FF74-645CFE37FB96 |
|
treatment provided by |
Plazi |
|
scientific name |
Sturnidoecus Eichler, 1944 |
| status |
|
Pediculus Linné, 1758 (in partim).
Philopterus Nitzsch, 1818: 288 (in partim). Docophorus Nitzsch, 1818: 289 (in partim). Nirmus Nitzsch, 1818: 291 (in partim).
Penenirmus Clay & Meinertzhagen, 1938: 73 (in partim). Sturnidoecus Eichler, 1944: 81 .
Turdinirmus Eichler, 1951b: 41 (in partim).
Type species. Docophorus leontodon Nitzsch, 1818: 290 [= Pediculus sturni Schrank, 1776: 118 ], by original designation.
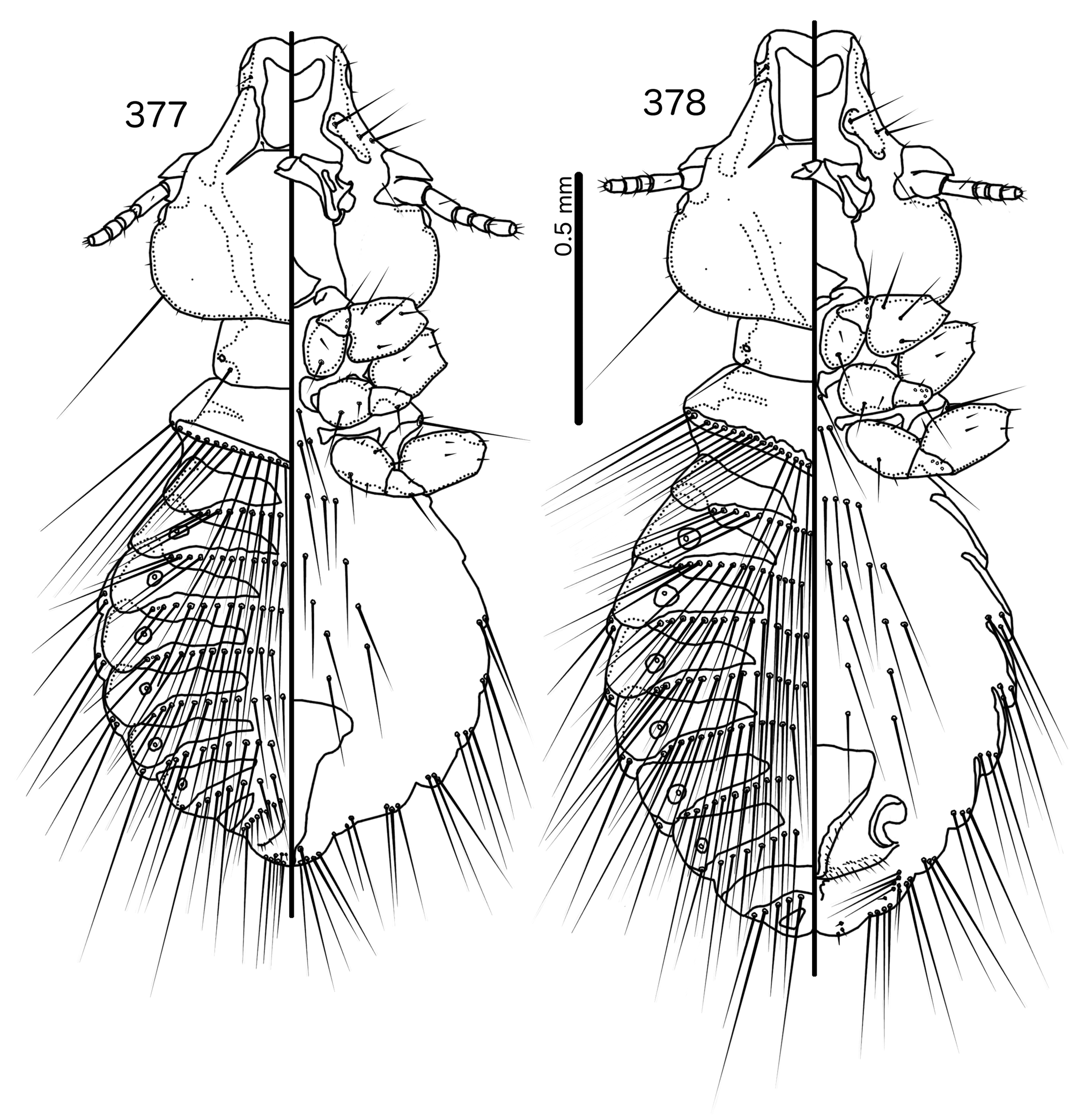
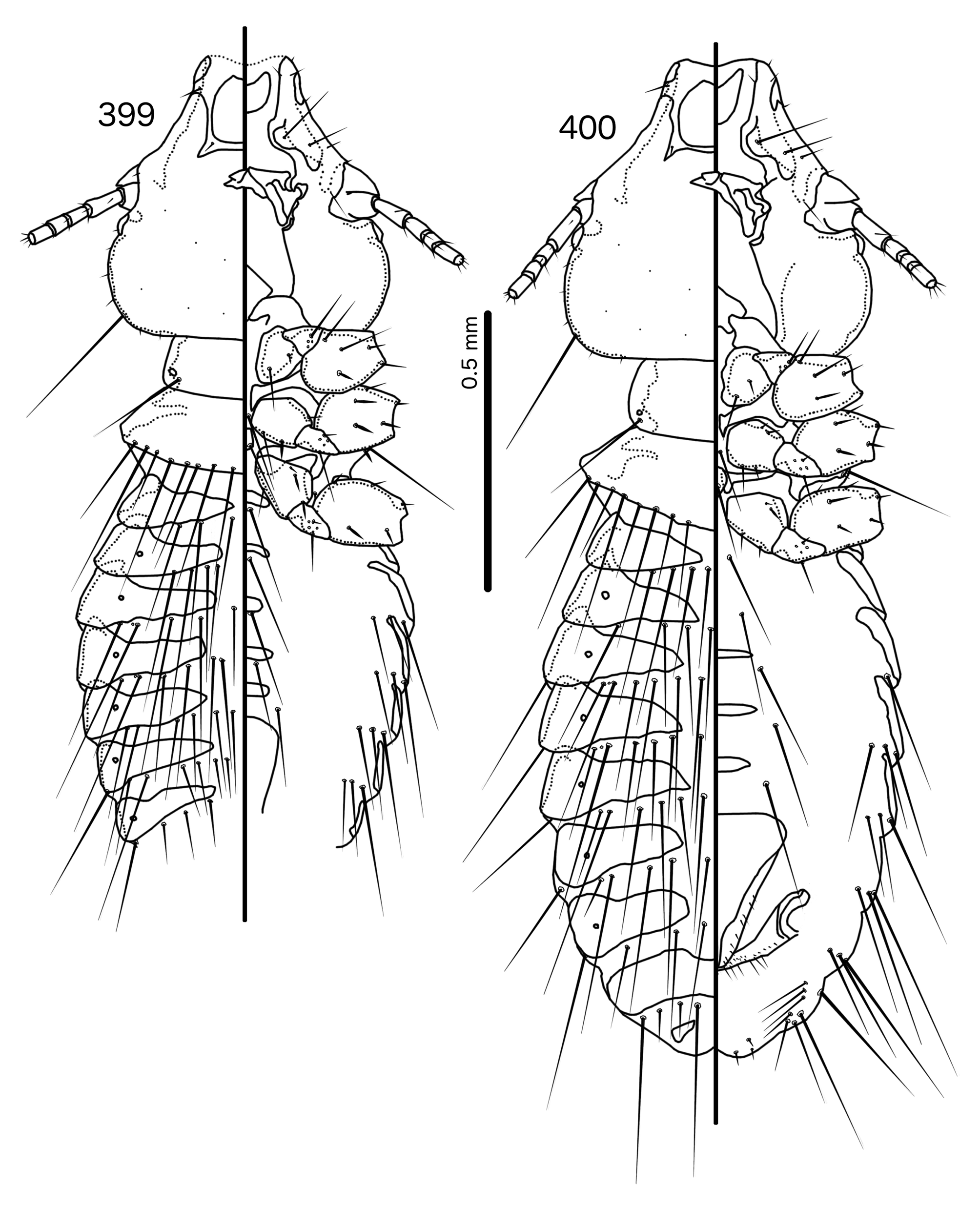
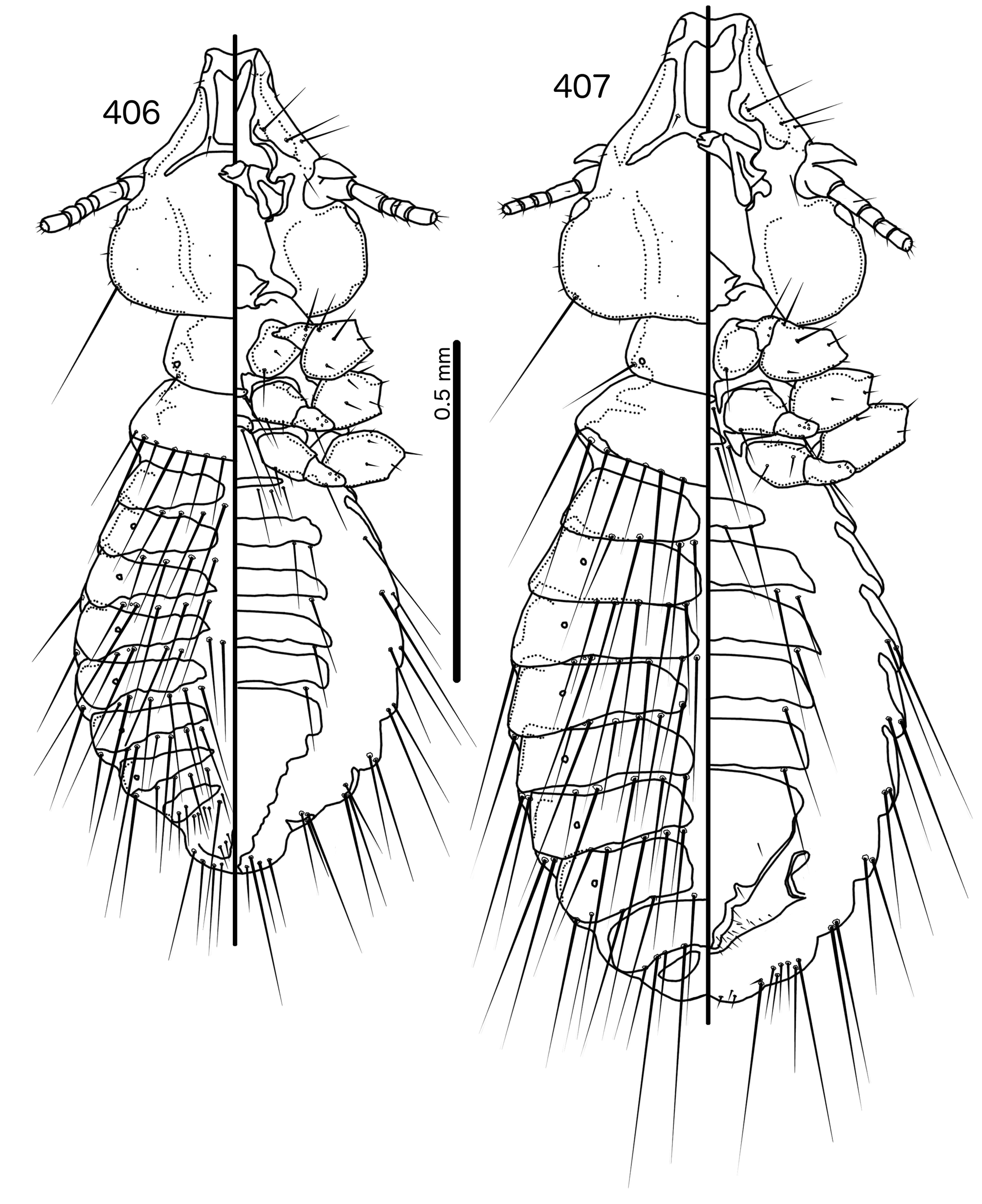
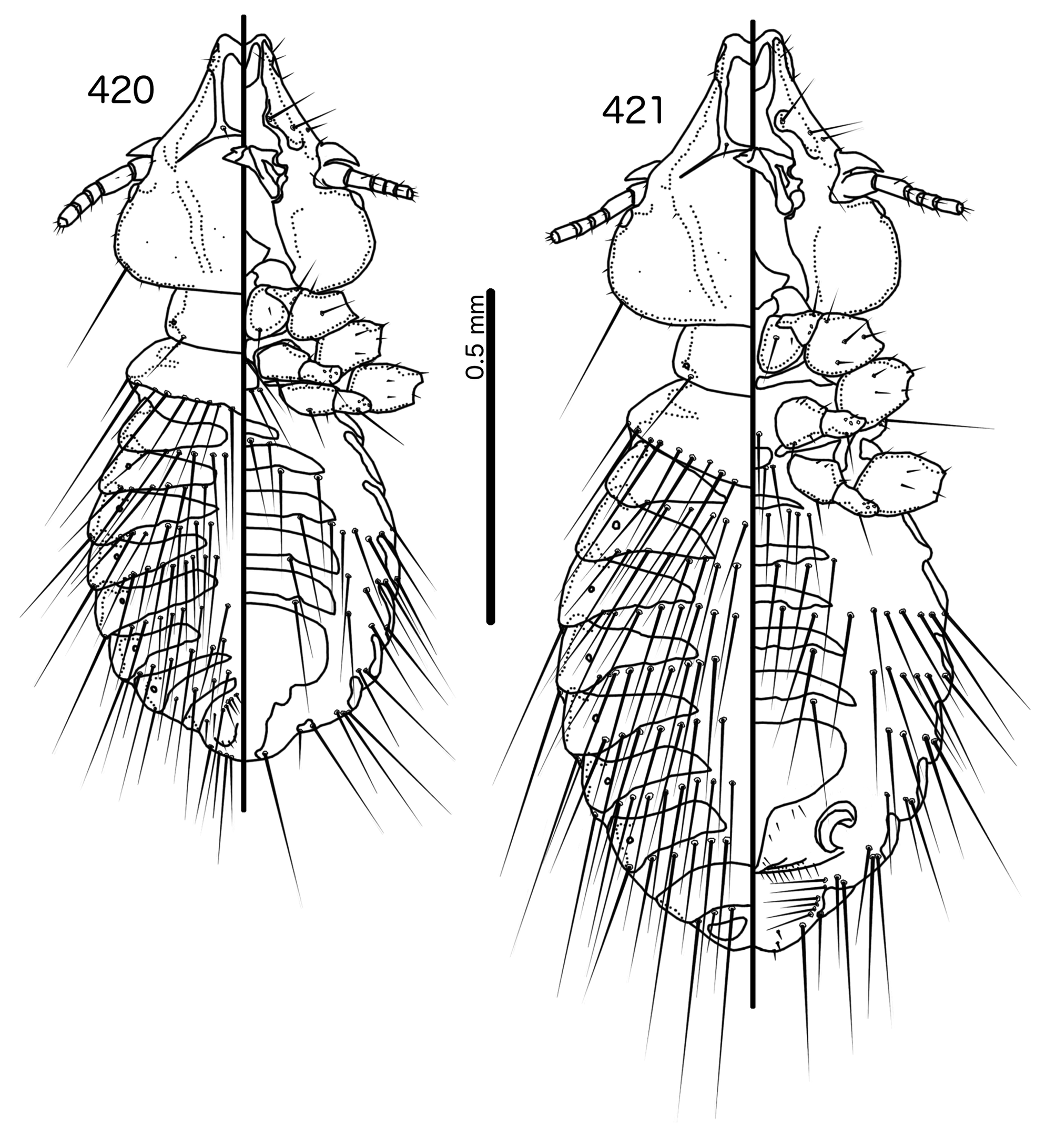
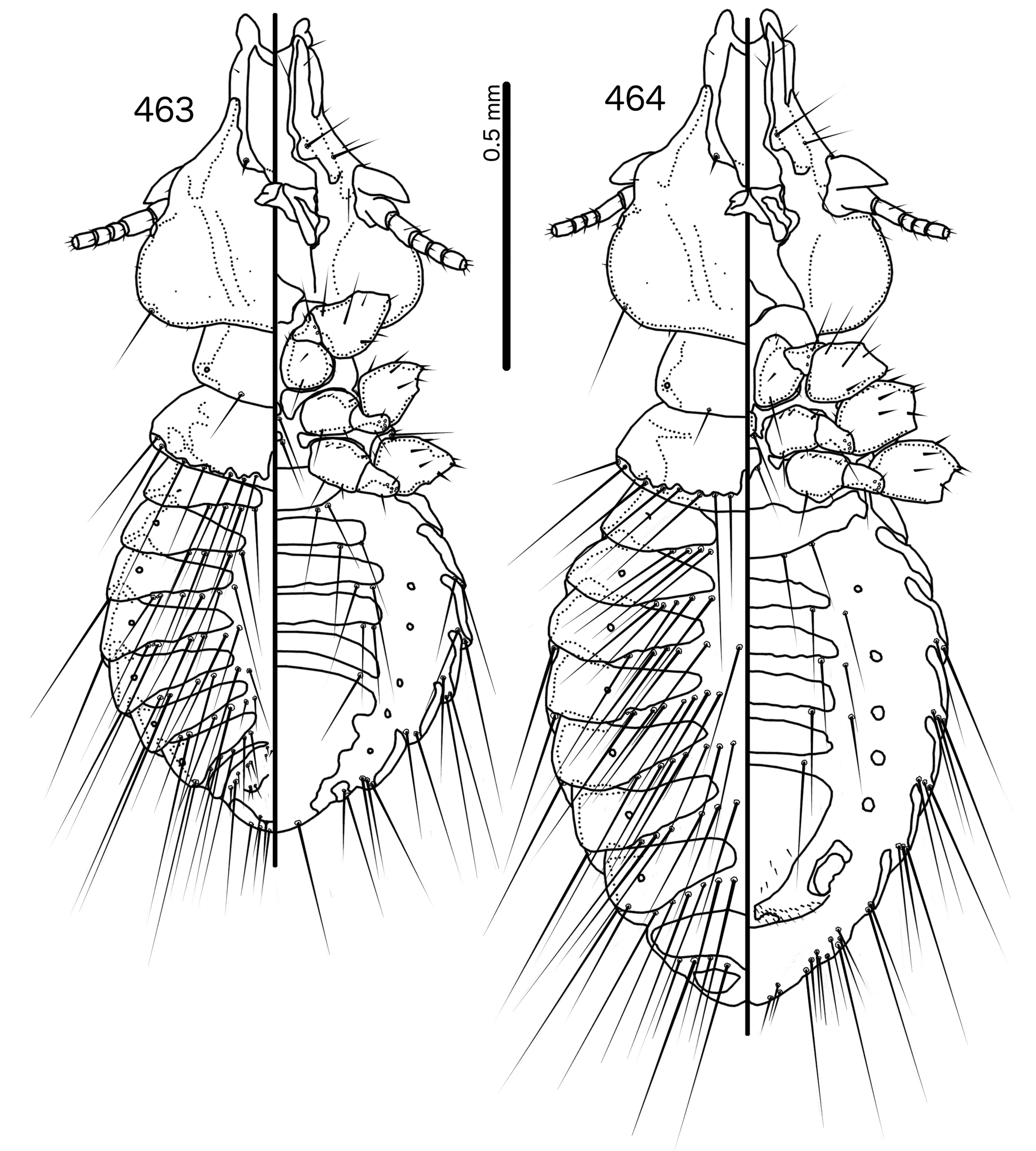
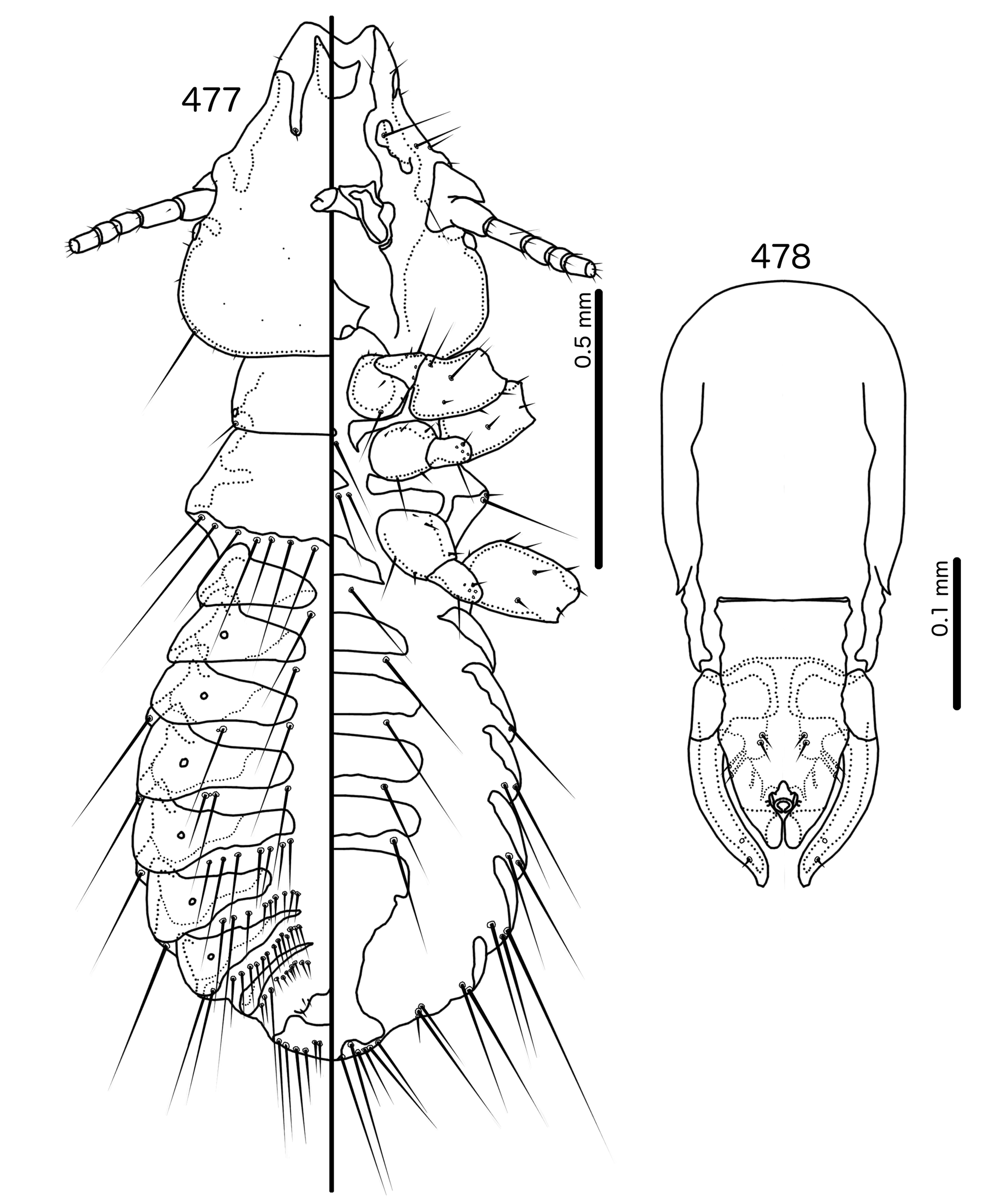
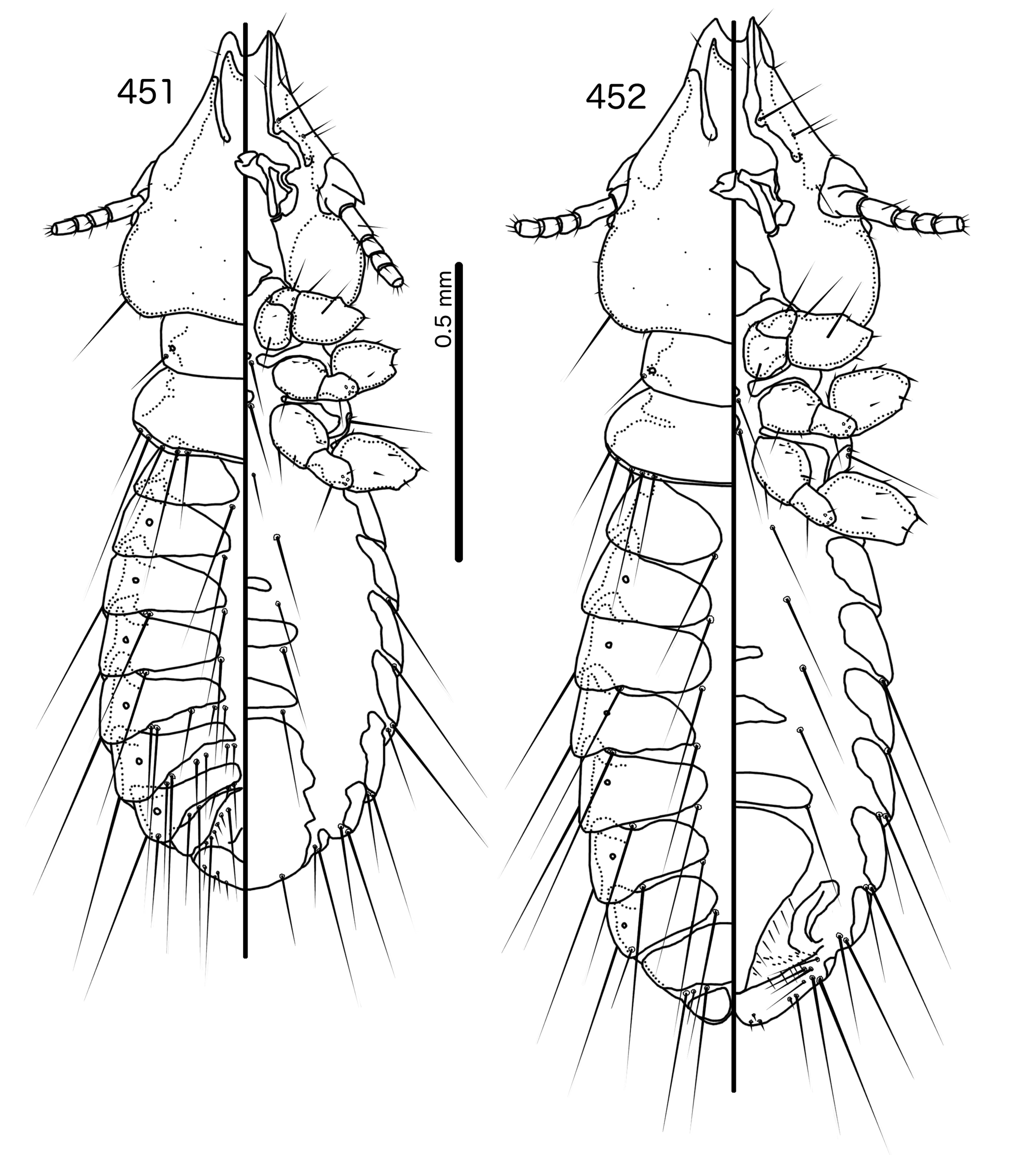
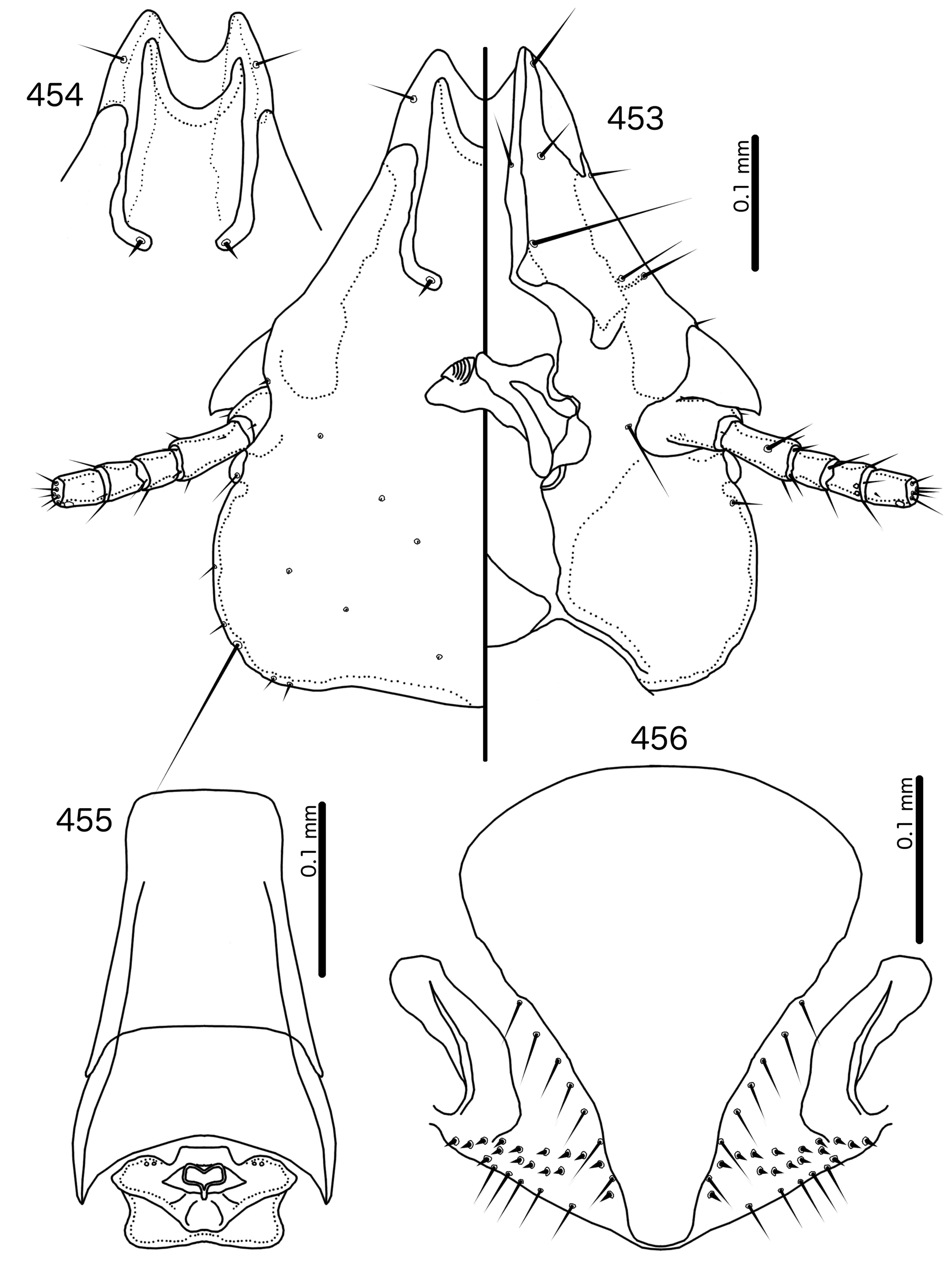
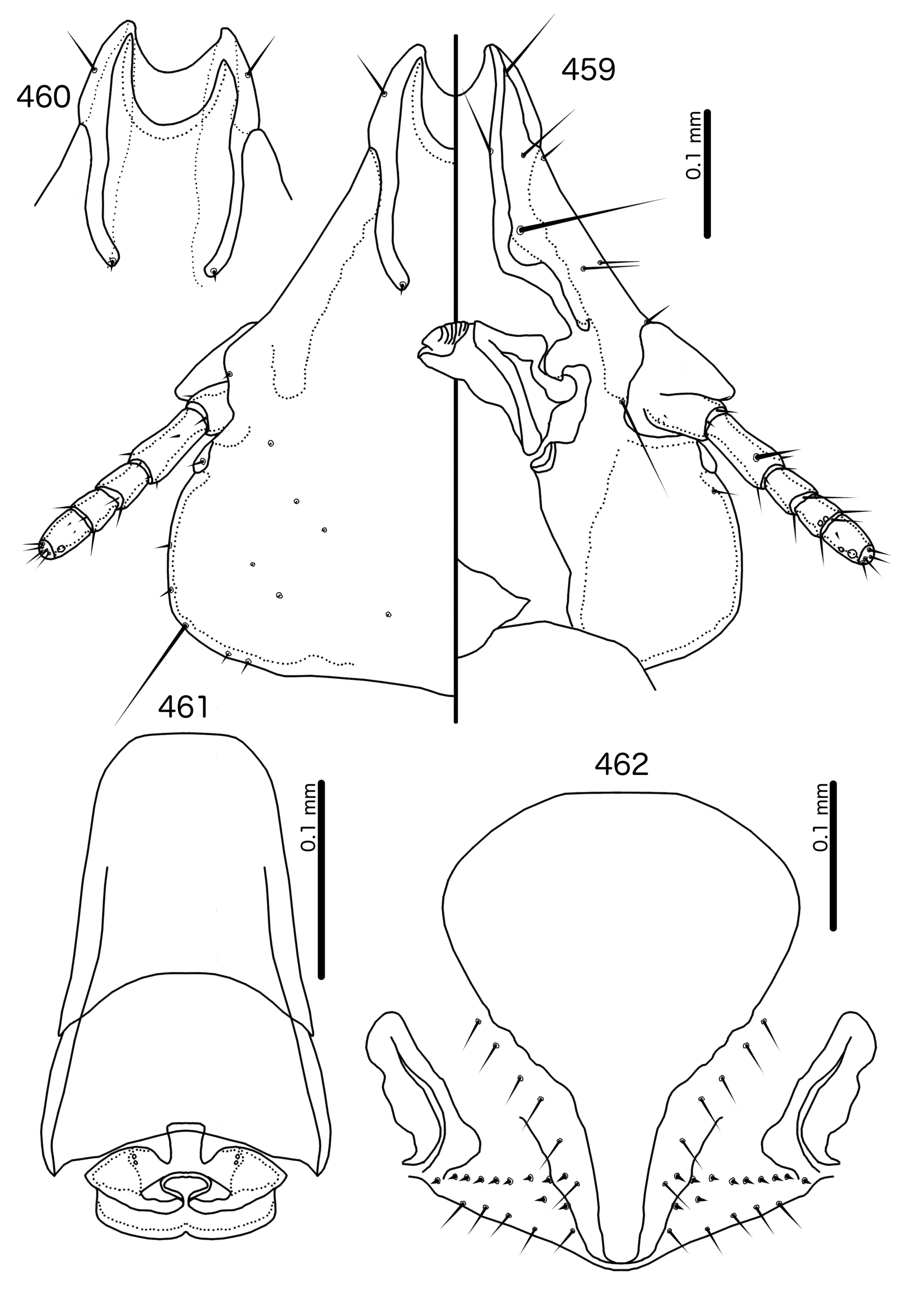
Diagnosis. In overall habitus, Sturnidoecus ( Figs 377–378 View FIGURES 377 – 378 , 399–400 View FIGURES 399 – 400 , 406–407 View FIGURES 406 – 407 , 413–414 View FIGURES 413 – 414 , 420–421 View FIGURES 420 – 421 ) is most similar to Rostrinirmus ( Figs 437–438 View FIGURES 437 – 438 ), Buphagoecus n. gen. ( Figs 427–428 View FIGURES 427 – 428 ), and Schizosairhynchus n. gen. ( Figs 463– 464 View FIGURES 463 – 464 , 470–471 View FIGURES 470 – 471 ). Members of all of these genera were considered parts of Sturnidoecus in the checklist of Price et al. (2003). Bizarrifrons ( Fig. 478 View FIGURES 477 – 478 ) and Manucodicola n. gen. ( Figs 451–452 View FIGURES 451 – 452 , 457–458) are also similar to Sturnidoecus , but these two genera are separated from Sturnidoecus species by the asymmetrical shape of the frons ( Figs 478 View FIGURES 477 – 478 , 454 View FIGURES 453 – 456 , 460 View FIGURES 459 – 462 ), and the structure of the dorsal anterior plate. In Bizarrifrons and Manucodicola , the dorsal anterior plate is continuous with the main head plate, whereas this plate is separate from the main head plate in Sturnidoecus .
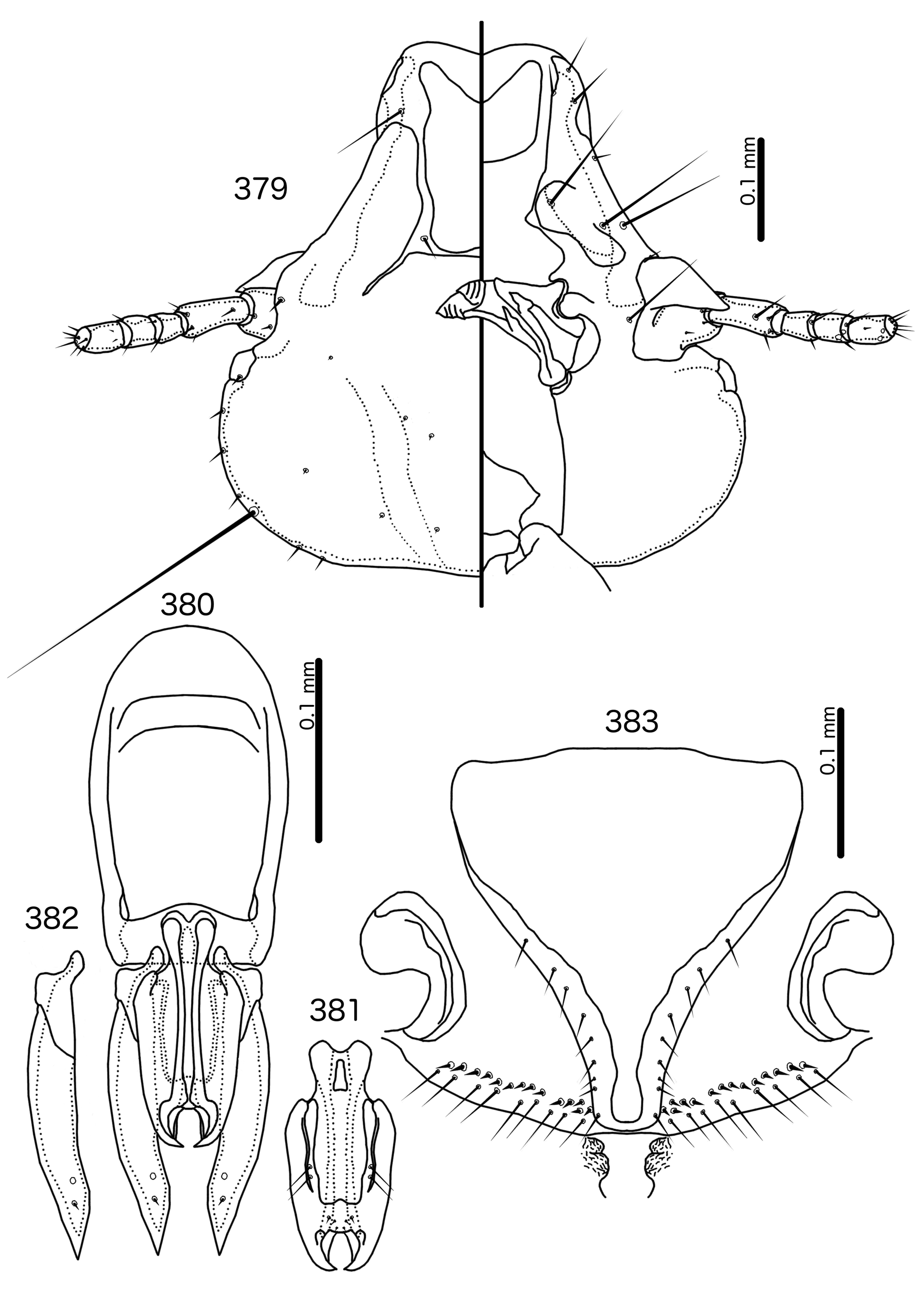
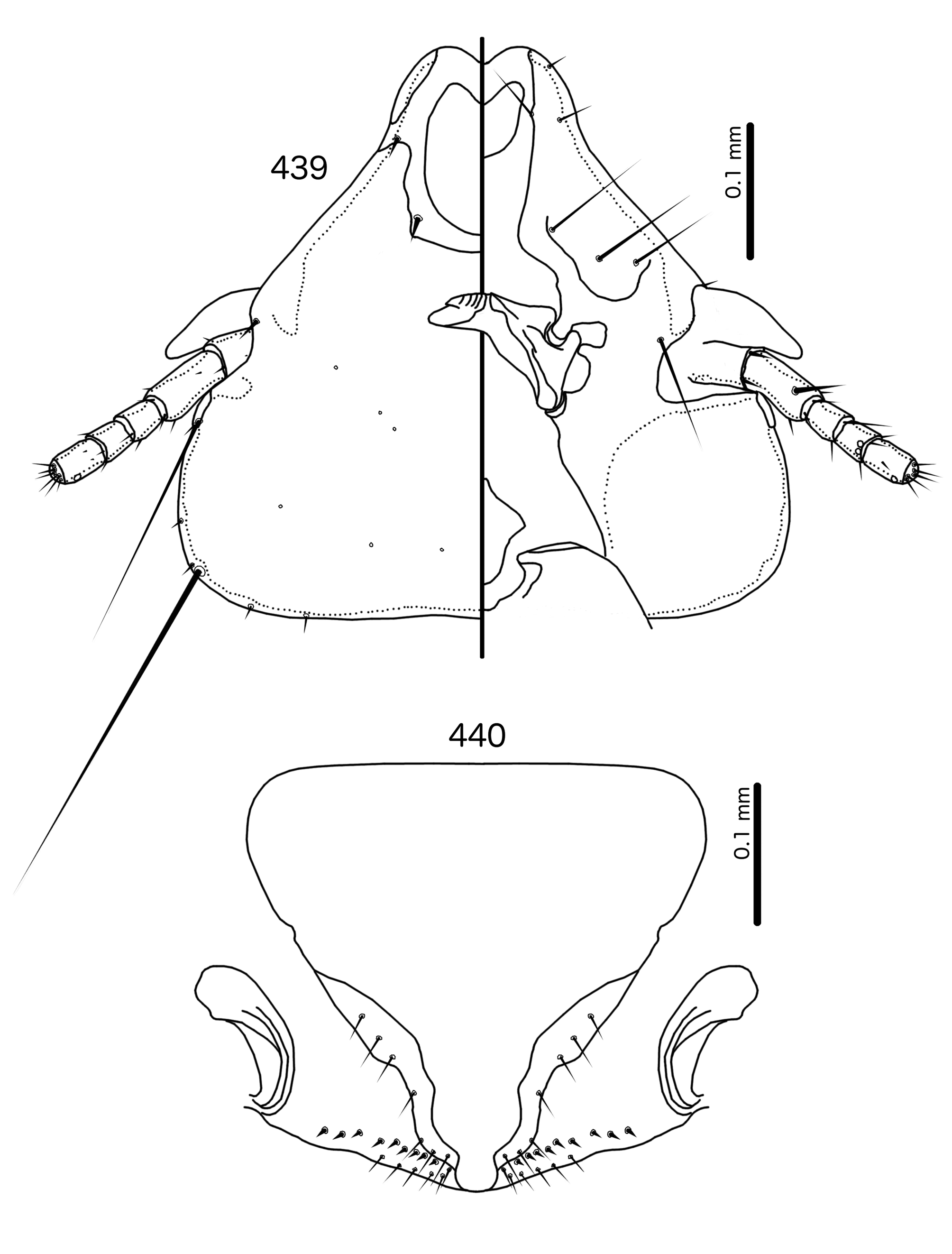
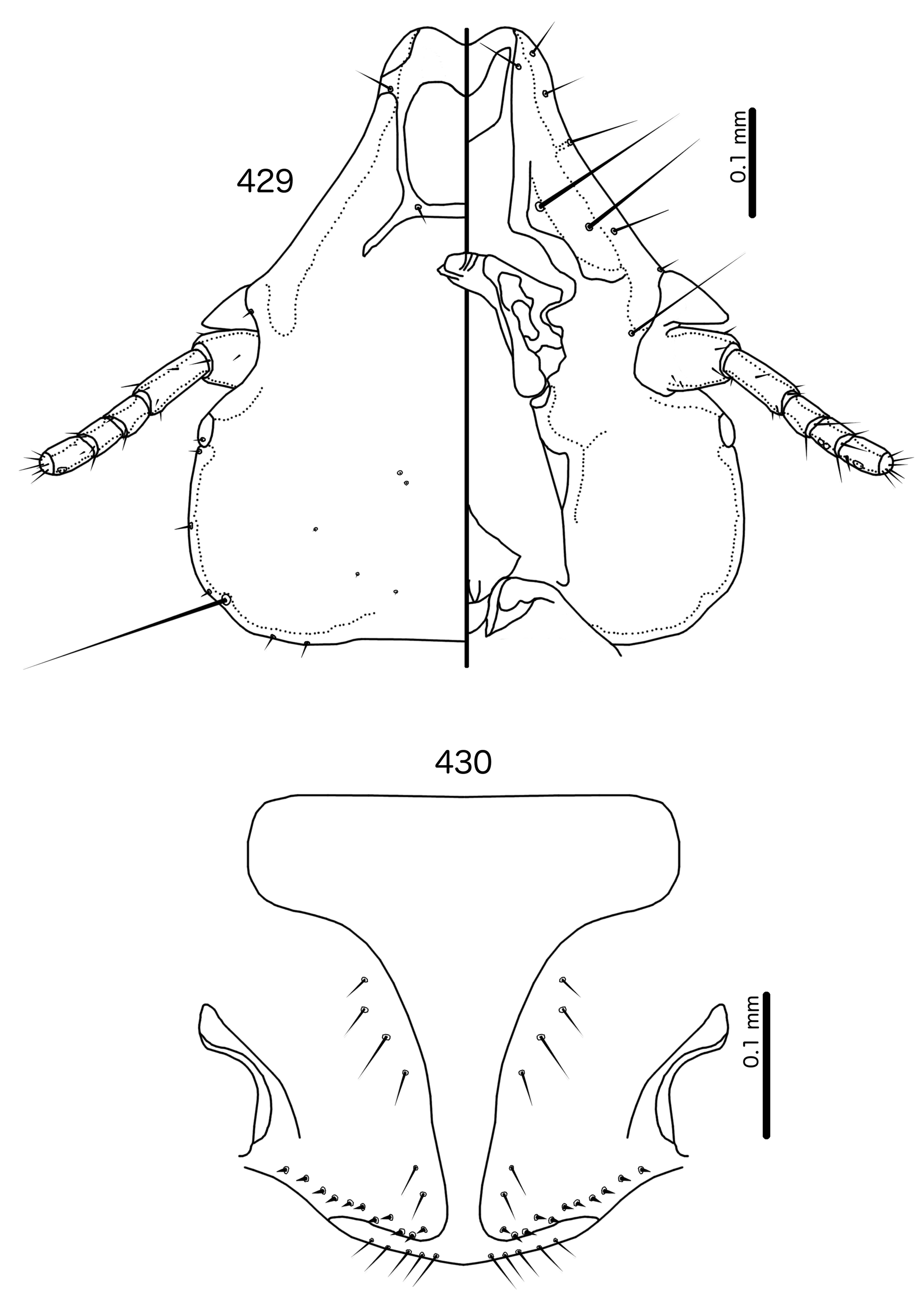
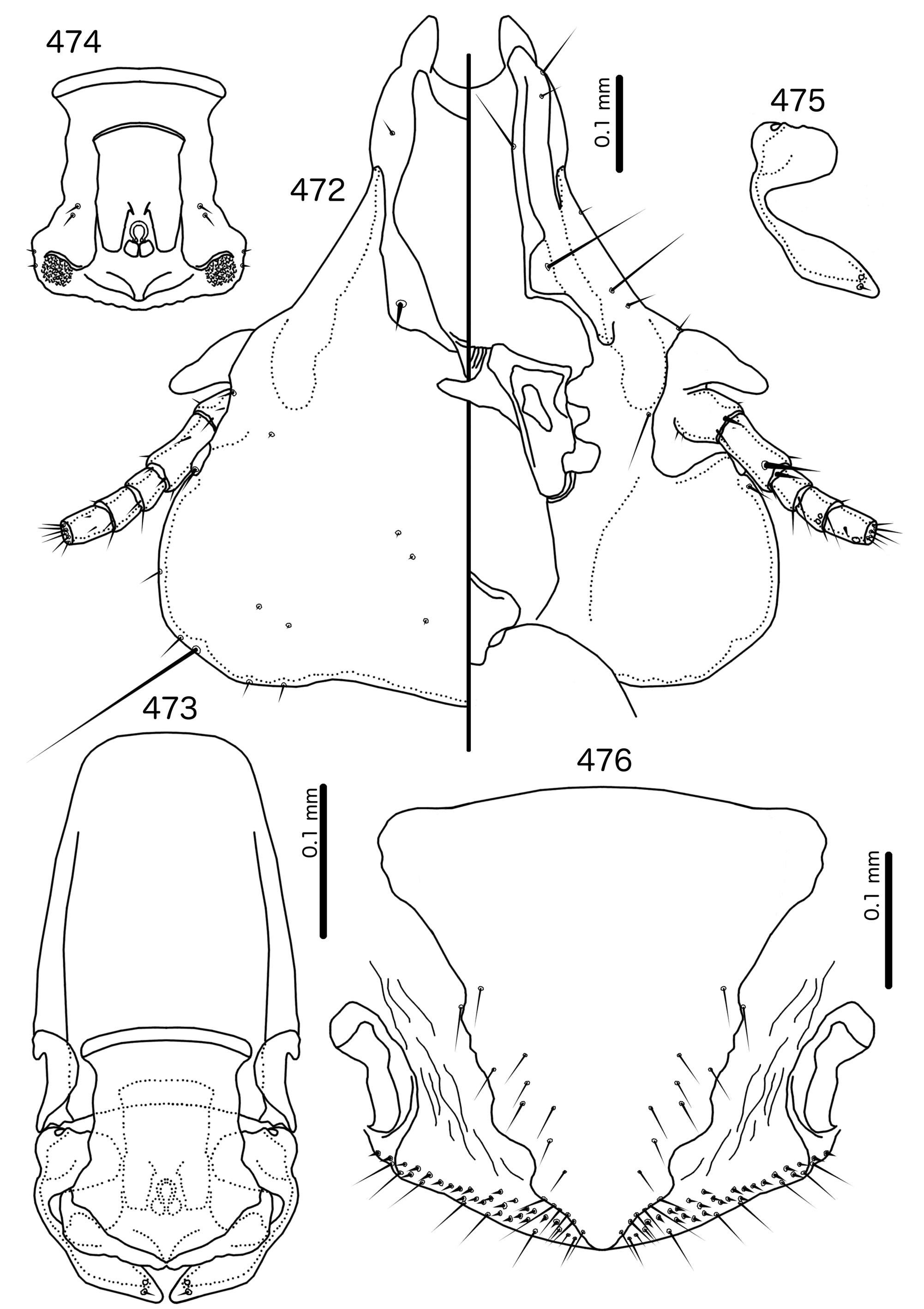
In Sturnidoecus ( Figs 379 View FIGURES 379 – 383 , 401 View FIGURES 401 – 405 , 408 View FIGURES 408 – 412 , 415 View FIGURES 415 – 419 , 422 View FIGURES 422 – 426 ), Rostrinirmus ( Fig. 439 View FIGURES 439 – 440 ), Buphagoecus ( Fig. 429 View FIGURES 429 – 430 ), and Schizosairhynchus ( Figs 465 View FIGURES 465 – 469 , 472 View FIGURES 472 – 476 ), the dorsal anterior plate is completely separated from the main head plate, and the dorsal preantennal suture reaches both the hyaline margin and the lateral margins of the head. Coni are large in all four genera, and typically reach to or beyond the distal margin of the scape. The marginal carina is interrupted laterally in all Sturnidoecus and Schizosairhynchus , but not in Rostrinirmus or Buphagoecus . The premarginal carina is always present in Sturnidoecus , but this is always absent in Schizosairhynchus . The postero-lateral extensions of the dorsal preantennal suture found in Sturnidoecus are also found in Buphagoecus , but not in Rostrinirmus or Schizosa. In all four genera as3 is absent. as1 is absent in Rostrinirmus , but this is present in Sturnidoecus . Conversely, as2 is absent in some Sturnidoecus (see species-group summaries below), but always present in the other three genera. The posterior margins of the dorsal anterior plate differs between these four genera, with those of Sturnidoecus and Buphagoecus roughly rectangular, those of Rostrinirmus more rounded, and those of Schizosairhynchus elongated into a thickened spur that overlaps with the main head plate.
Sturnidoecus ( Figs 377–378 View FIGURES 377 – 378 , 399–400 View FIGURES 399 – 400 , 406–407 View FIGURES 406 – 407 , 413–414 View FIGURES 413 – 414 , 420–421 View FIGURES 420 – 421 ) and Schizosairhynchus ( Figs 463–464 View FIGURES 463 – 464 , 470–471 View FIGURES 470 – 471 ) have setal rows on most tergopleurites. By contrast, in Buphagoecus ( Figs 427–428 View FIGURES 427 – 428 ) and Rostrinirmus ( Figs 437–438 View FIGURES 437 – 438 ) all tergopleurites of both sexes have at most 1 ss, 1 psps, and 1 aps , apart from ps. Rostrinirmus, Schizosairhynchus , and Sturnidoecus all have multiple sts on at least some segments, but this is never found in Buphagoecus . No Sturnidoecus have subsidiary lateral sternal plates or the wide sternal plate II found in all Schizosairhynchus .
The female genitalia are virtually the same in both Sturnidoecus ( Figs 383 View FIGURES 379 – 383 , 405 View FIGURES 401 – 405 , 412 View FIGURES 408 – 412 , 419 View FIGURES 415 – 419 , 426 View FIGURES 422 – 426 ) and Rostrinirmus ( Fig. 440 View FIGURES 439 – 440 ), but many Schizosairhynchus ( Fig. 469 View FIGURES 465 – 469 ) and Buphagoecus ( Fig. 430 View FIGURES 429 – 430 ) have partial crosspieces. In general, females of Sturnidoecus and Schizosairhynchus have more vms than females of the other two genera, but there is considerable intraspecific variation in setal numbers.
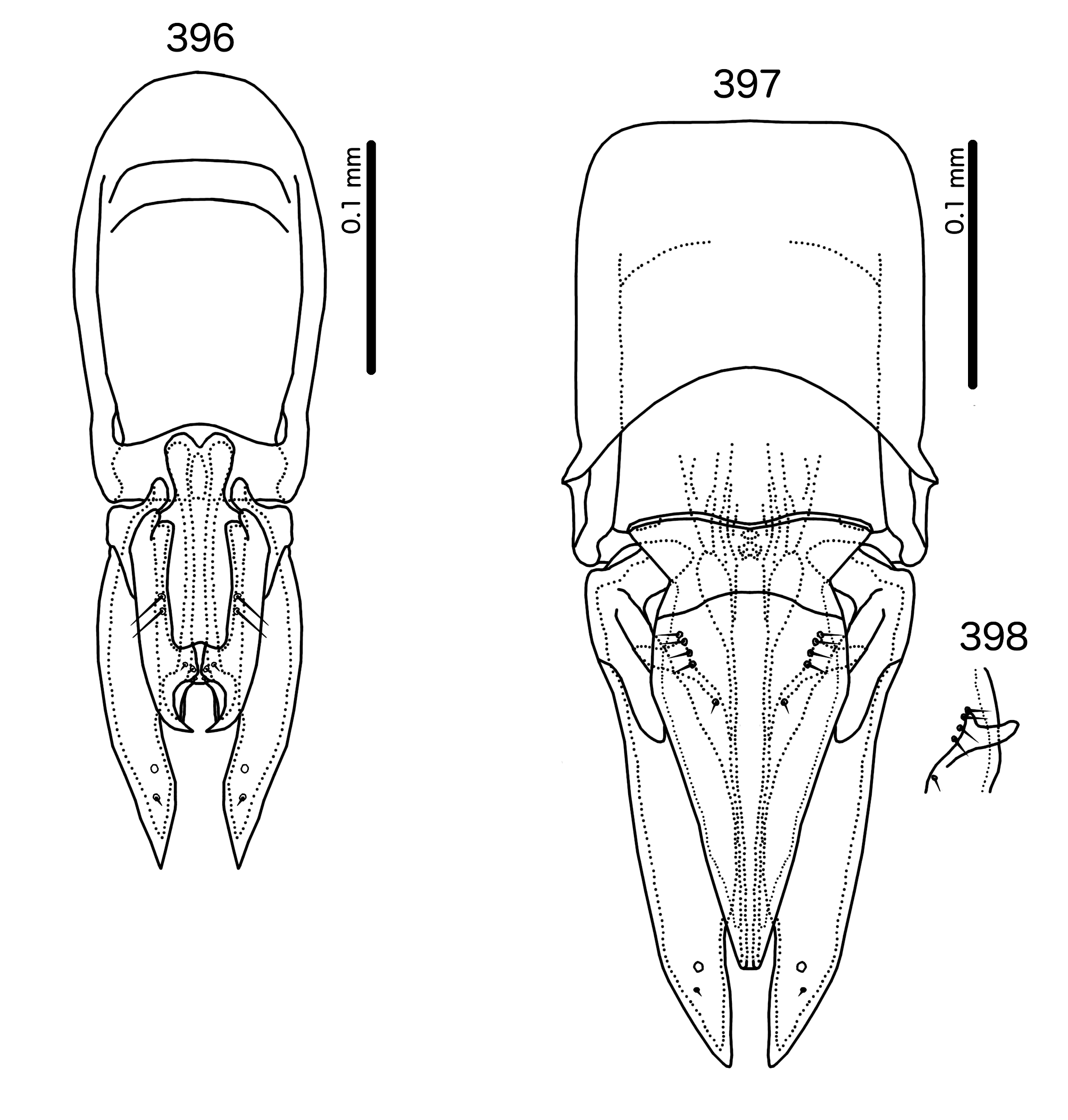
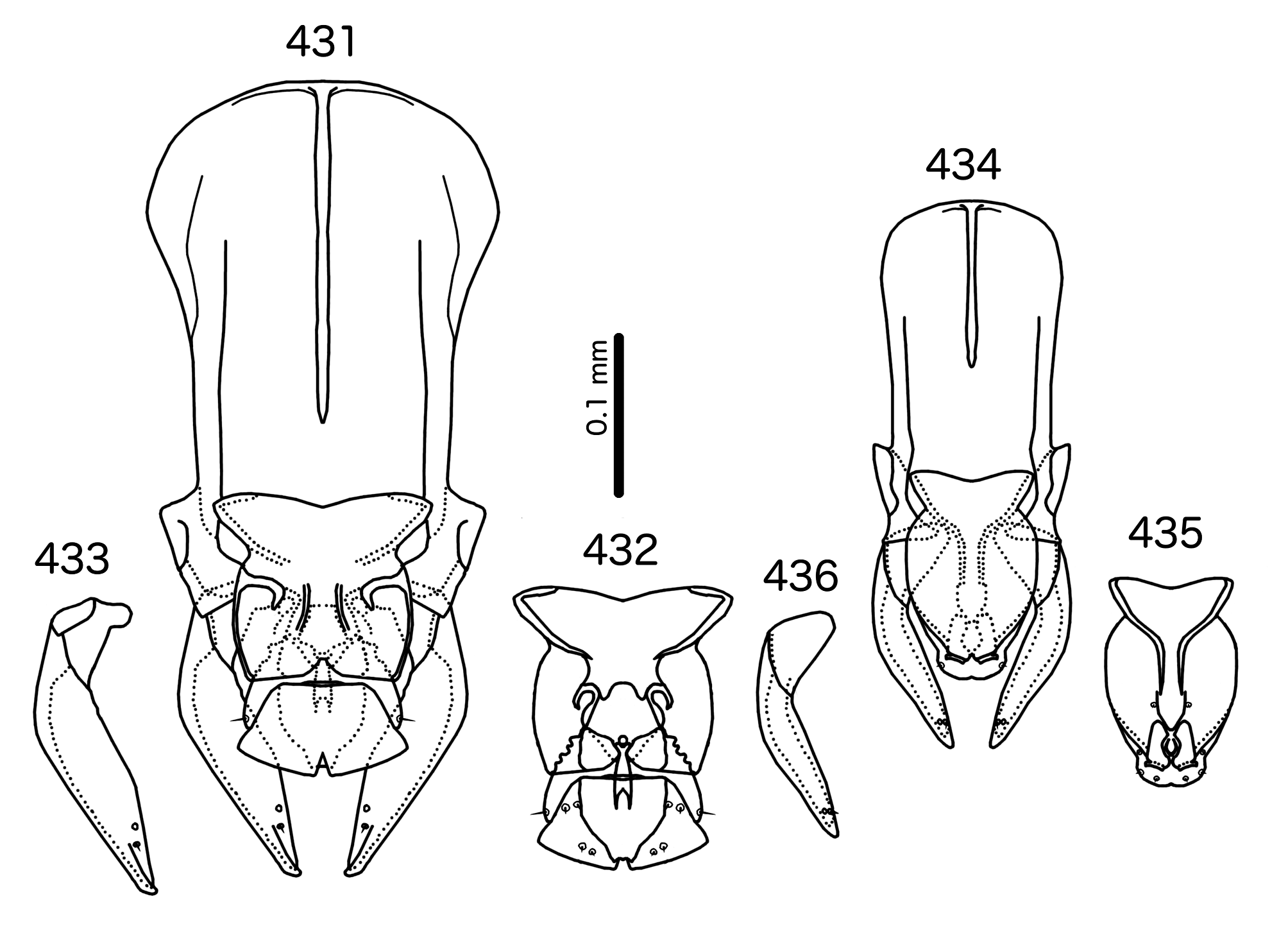
The male genitalia of Sturnidoecus are variable ( Figs 384–398 View FIGURES 384 – 389 View FIGURES 390 – 395 View FIGURES 396 – 398 ), but distinctly different from those of Buphagoecus ( Figs 431–436 View FIGURES 431 – 436 ), Rostrinirmus ( Figs 441–443, 445–450 View FIGURES 441 – 450 ), and Schizosairhynchus ( Figs 466–468 View FIGURES 465 – 469 , 474– 476 View FIGURES 472 – 476 ). The gonopore is always terminal in Sturnidoecus (but not clearly visible in St. somnodraco n. sp., Figs 423– 425 View FIGURES 422 – 426 ), but ventral in Schizosairhynchus , Rostrinirmus , and Buphagoecus . Unlike in Schizosairhynchus and Rostrinirmus , there are no rugose nodi in any Sturnidoecus , and the hook-shaped or triangular ventral processes found in many Sturnidoecus groups are not found in any other genus in the Brueelia -complex. While mesosomal setal patterns vary considerably among the different species-groups in Sturnidoecus , there are a few setal characters that set Sturnidoecus spp. apart: the pmes are never on the lateral margins of the mesosome in Sturnidoecus , but they are always on the lateral margins in Schizosairhynchus ; in Buphagoecus , both pmes and ames may be distal to gonopore (e.g. Fig. 432 View FIGURES 431 – 436 ), but this is never the case in Sturnidoecus .
Outside the Brueelia -complex, Sturnidoecus is superficially similar to the Philopterus - and Penenirmus - complexes, which occasionally infest the same hosts as Sturnidoecus spp. (see Price et al. 2003). All three of these generic complexes contain species that are of the “head louse” ecotype ( Johnson et al. 2012), and at least some species of all three generic complexes have the following characters in common: marginal carina interrupted submedianly and laterally; dorsal anterior plate completely separated from main head plate posteriorly; at least some tergopleurites with dense or sparse setal rows; pleurites that do not, or only barely, reach the ventral surface of the abdomen. However, there are also substantial differences between Sturnidoecus and members of the Philopterus - and Penenirmus -complexes. The hyaline margin in Sturnidoecus is always contained between the antero-lateral ends of the marginal carina, and the laterally extended hyaline frons seen in some Philopterus - complex genera [e.g. Mayriphilopterus Mey 2004 (see Mey 2004: fig. 17a)] does not occur in Sturnidoecus . The posterior margin of the dorsal anterior plate is roughly flat in all Sturnidoecus (e.g. Fig. 379 View FIGURES 379 – 383 ), not tapered posteriorly as in many Philopterus -complex genera, nor pointed as in the Penenirmus -complex. The mts 3 is the only temporal macrosetae in Sturnidoecus ( Fig. 379 View FIGURES 379 – 383 ), and neither os nor pos are macrosetae in this genus, unlike in the Penenirmus - and Philopterus -complexes. Trabecula are absent in the Penenirmus -complex and in Sturnidoecus ( Fig. 379 View FIGURES 379 – 383 ), but present in the Philopterus -complex; however, the long and often thick coni of Sturnidoecus may be confused with trabecula, especially in poorly preserved material or nymphs. In the Philopterus -complex ads is located on the posterior margin of the dorsal anterior plate, whereas in Sturnidoecus ( Fig. 379 View FIGURES 379 – 383 ) it is always located in the dorsal preantennal suture. Additional differences between lice in the Penenirmus - and Philopterus - complexes and lice of the Brueelia-complex are detailed in the introduction.
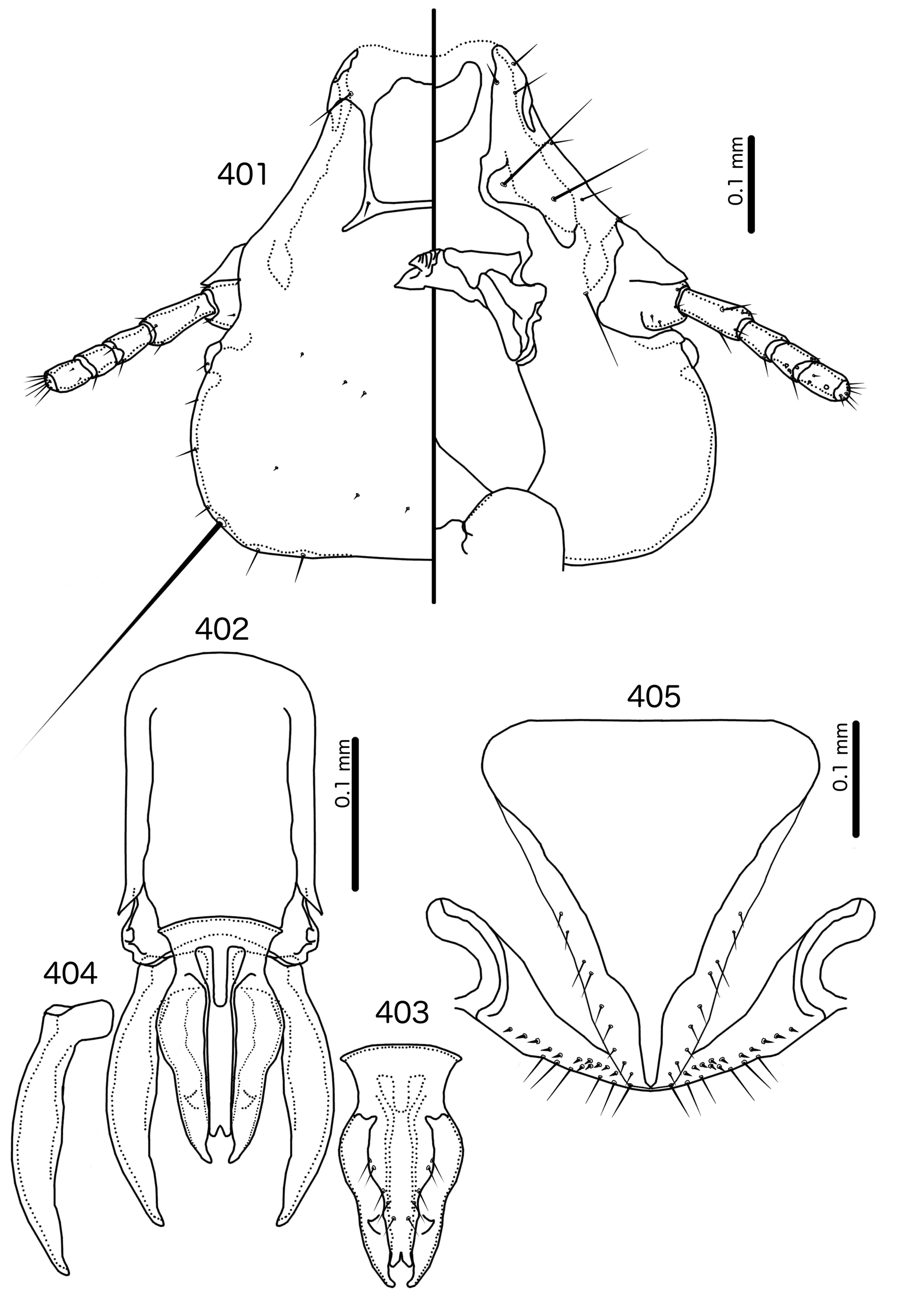
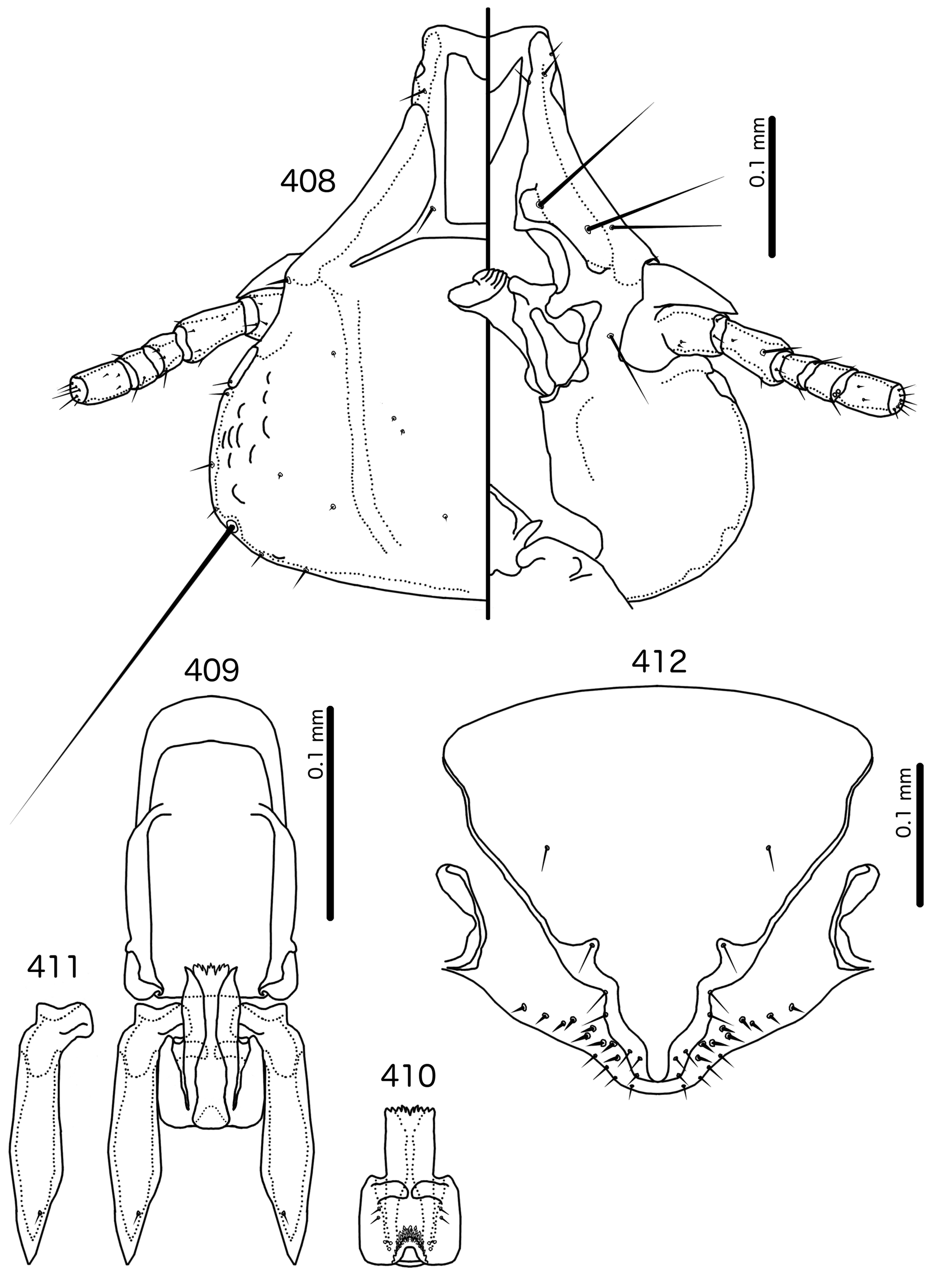
Description. Both sexes. Head bulb-shaped ( Figs 379 View FIGURES 379 – 383 , 401 View FIGURES 401 – 405 , 408 View FIGURES 408 – 412 , 415 View FIGURES 415 – 419 , 422 View FIGURES 422 – 426 ). Marginal carina interrupted submedianly, but typically entire laterally; however, the lateral section of the margila carina where the dorsal preantennal suture reaches the lateral margin of the head is typically indented laterally ( Fig. 415 View FIGURES 415 – 419 ) or displaced medianly ( Fig. 379 View FIGURES 379 – 383 ). Frons flat or concave, hyaline. Hyaline margin continuous with dorsal preantennal suture reaching dsms and ads, completely separating dorsal anterior plate from main head plate. Postero-lateral corners of suture extended towards preantennal nodi. Dorsal anterior plate with flat or concave posterior margin, concave anterior margin. Ventral anterior plate present, crescent-shaped. Ventral carinae diffuse anterior to pulvinus. Head setae as in Figs 379 View FIGURES 379 – 383 , 401 View FIGURES 401 – 405 , 408 View FIGURES 408 – 412 , 415 View FIGURES 415 – 419 , 422 View FIGURES 422 – 426 , varying slightly between species groups; as3 absent; as1 absent in Sturnidoecus mon n. sp. ( Fig. 408 View FIGURES 408 – 412 ) and St. somnodraco n. sp. ( Fig. 422 View FIGURES 422 – 426 ) species-groups. Coni long, pointed, often reaching beyond distal margin of scapes. Antennae monomorphic (except St. orientalis ; not illustrated). Temporal carinae visible in most species; mts 3 only macrosetae. Gular plate variable.
Prothorax ( Figs 377–378 View FIGURES 377 – 378 , 399–400 View FIGURES 399 – 400 , 406–407 View FIGURES 406 – 407 , 413–414 View FIGURES 413 – 414 , 420–421 View FIGURES 420 – 421 ) small, rectangular; ppss on postero-lateral corners. Proepimera hook- or hammer-shaped medianly. Pterothorax trapezoidal to pentagonal; lateral margins divergent; posterior margin either convergent to median point, flat, or gently rounded; mms uninterrupted medianly. Meso- and metasterna small, not fused. Mesosternum with 1 seta on postero-lateral corner on each side; metasternum with 1–3 setae on postero-lateral corner on each side. Metepisterna with slender to hammer-shaped median ends. Leg chaetotaxy as in Fig. 25 View FIGURES 25 , except fI-p2 absent.
Abdomen ( Figs 377–378 View FIGURES 377 – 378 , 399–400 View FIGURES 399 – 400 , 406–407 View FIGURES 406 – 407 , 413–414 View FIGURES 413 – 414 , 420–421 View FIGURES 420 – 421 ) round to oblong in female, rounded in males; almost circular in some species. Tergopleurites rectangular or triangular; tergopleurites II–IX+X in male and tergopleurites II–VIII in female moderately to widely separated medianly. Sternal plates medianly continuous, variable in shape between species groups, and some or all may be absent. Pleural incrassations slight; ventral segments of the pleurites typically absent at least in more posterior segments. Re-entrant heads broad, blunt. Abdominal chaetotaxy as in Table 2, but differs between species groups ( Table 11). Male subgenital plate triangular to trapezoidal, in some species may not reach posterior margin of abdomen. Female subgenital plate ( Figs 383 View FIGURES 379 – 383 , 405 View FIGURES 401 – 405 , 412 View FIGURES 408 – 412 , 419 View FIGURES 415 – 419 , 426 View FIGURES 422 – 426 ) roughly triangular, lateral margins often concave, approaching or reaching vulval margin; cross-piece or lateral extensions never present. Vulval margin ( Figs 383 View FIGURES 379 – 383 , 405 View FIGURES 401 – 405 , 412 View FIGURES 408 – 412 , 419 View FIGURES 415 – 419 , 426 View FIGURES 422 – 426 ) with slender vms, thorn-like vss; vos follows lateral margins of subgenital plate; distal vos typically median to vss.
Species Sex ps aps psps tps ss sts
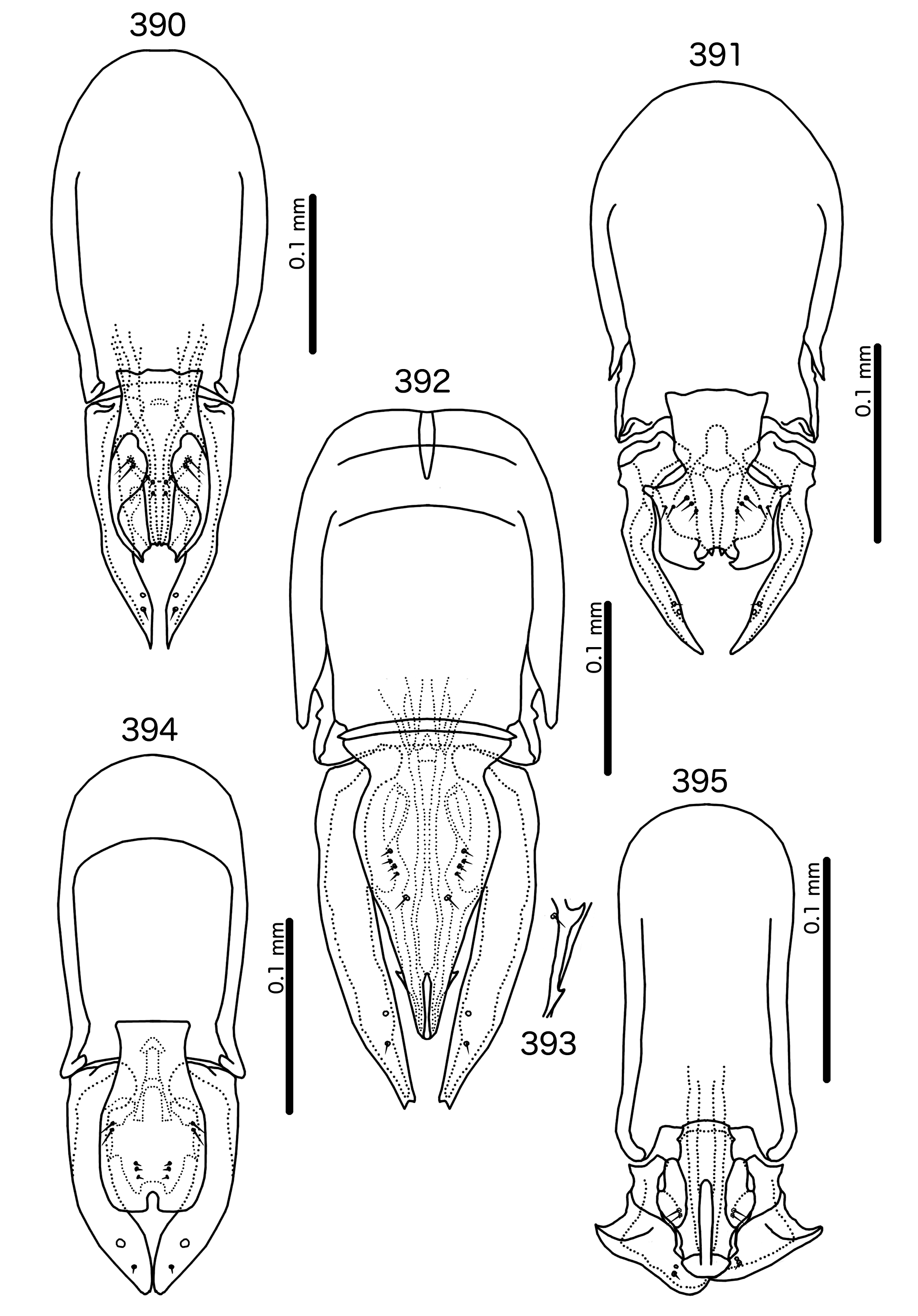
St. sturni M IV–VIII II–VII II–VIII II–VIII II–VIII II–VI F IV–VIII II–VII II–VIII II–VIII II–VIII II–VI St. australafricanus M IV–VIII IV–VII IV–VIII II–VIII II–VIII II–VI F IV–VIII IV–VII III–VIII II–VIII II–VIII II–VI St. mon M III–VIII IV–VII IV–VIII II–VIII II–VIII II–VI F IV–VIII IV–VII IV–VIII II–VIII II–VIII II–VI St. porphyrogenitus M IV–VIII V–VII IV–VIII II–VIII II–VIII II–VI F IV–VIII VI–VII IV–VIII II–VIII II–VIII II–VI Male genitalia differ between species-groups ( Figs 384–398 View FIGURES 384 – 389 View FIGURES 390 – 395 View FIGURES 396 – 398 ). Basal apodeme generally broad, rounded to rectangular. Proximal mesosome typically broad. Gonopore highly modified, and often not recognizable; terminal in all species-groups except Sturnidoecus somnodraco ( Fig. 395 View FIGURES 390 – 395 ). Mesosomal lobes usually very slender, may be elongated distally. Mesosome may be much shorter than parameres (St. mon species-group, Fig. 389 View FIGURES 384 – 389 ) to almost as long as parameres ( St. clayae species-group, Fig. 386 View FIGURES 384 – 389 ). Up to 5 setae on each side of mesosome, location varies between species groups, but typically separated into longer ames and shorter pmes; pmes typically more median than ames. Several groups (e.g. St. clayae species group, Fig. 387 View FIGURES 384 – 389 ) with one or more fleshy triangular structures on ventral surface. Parameral heads folded, typically rectangular (e.g. St. simpl ex species-group, Fig. 394 View FIGURES 390 – 395 ) or fingerlike (e.g. St. meinertzhageni species-group, Fig. 388 View FIGURES 384 – 389 ), but may be different (e.g. St. clayae species group, Fig. 386 View FIGURES 384 – 389 ). Parameral blades typically slender, elongated (but see St. somnodraco species-group, Fig. 395 View FIGURES 390 – 395 ); pst1 sensilla, typically central; pst2 microsetae, central or submarginal near median margin of parameres.
Species-groups. All species groups in Sturnidoecus are based on the structure of the male genitalia. Representative male genitalia for each species group are illustrated in Figs 384–398 View FIGURES 384 – 389 View FIGURES 390 – 395 View FIGURES 396 – 398 . Note that these species groups are only for the convenience of identification and comparison, and may not from monophyletic groups. The phylogeny of Bush et al. (2016) did not represent all of the groups with distinct male genitalia, and therefore how these unique genitalic structures are distributed across Sturnidoecus needs further study.
Material from several species were not available to us, and the existing descriptions and illustrations are insufficiently detailed to assign these species of the groups used here. Thus, these species are presently placed as Sturnidoecus incertae sedis (see below). The single female of St. femoratus ( Piaget, 1880) examined by us also cannot be placed. Note also that some species listed as Sturnidoecus by Price et al. (2003) are here removed to Penenirmus or Philopterus . These are principally species described by Ansari (1955b), that have too many temporal macrosetae, and in some cases trabecula, as is apparent in the later illustrations provided by Ansari (1958b). These species are discussed towards the end of this paper, under “Species referred to other genera”, and include: Docophorus aenas Piaget, 1885 , Sturnidoecus capitis Ansari, 1955b , and Sturnidoecus chilchil Ansari, 1955b , which are moved to Philopterus ; and Sturnidoecus chendoola Ansari, 1955b , Sturnidoecus guldum Ansari, 1955b , Sturnidoecus irritans Ansari, 1955b , Nirmus quadrilineatus Nitzsch, 1866 , and Sturnidoecus saleimi Ansari, 1955b , which are moved to Penenirmus .
In addition, we have transferred Sturnidoecus philippensis Tandan & Kumar, 1969 , to the genus Schizosairhynchus (see above), and St. husaini Ansari, 1968 , and St. prominens Ansari, 1968 , have been elevated to a genus of their own, Buphagoecus .
Sturnidoecus australafricanus species-group ( Fig. 384 View FIGURES 384 – 389 ). as1 present. Basal apodeme rectangular, anterior end rounded. Proximal mesosome as in Fig. 384 View FIGURES 384 – 389 , wider anteriorly, overlapping basal apodeme. Mesosomal lobes narrow, sinuous, with wide lateral thickenings; ames and pmes of roughly equal length, barely separated into sets, and together forming distally converging rows. Ventral triangular hook just distal to midline. Parameral heads rectangular. Parameral blades gently tapering, curving medianly.
Sturnidoecus basilewskyi species-group ( Fig. 385 View FIGURES 384 – 389 ). as1 present. Basal apodeme rectangular, anterior end rounded. Proximal mesosome as in Fig. 385 View FIGURES 384 – 389 , wider anteriorly, overlapping basal apodeme. Mesosomal lobes slender, thickened laterally, with prominent combs distally; 2 ames on each side much longer than 3 pmes on each side; ames and pmes aligned to form distally converging rows. Parameral heads slender. Parameral blades slender, with irregular median margin.
Sturnidoecus clayae species-group ( Figs 386–387 View FIGURES 384 – 389 ). as1 present. Basal apodeme truncated oval, anterior end rounded. Proximal mesosome as in Fig. 386 View FIGURES 384 – 389 , wider anteriorly, not overlapping basal apodeme. Mesosomal lobes slender, elongated to reach distal tips of parameres. Elongated parts of mesosome not fused medianly. Triangular extension on ventral side ( Fig. 387 View FIGURES 384 – 389 ); 3 ames much longer than 2 pmes; ames sublateral, not forming distally converging rows; pmes lateral to gonopore, forming transversal row. Parameral heads wider medianly. Parameral blades elongated, straight, pointed distally.
Sturnidoecus meinertzhageni species-group ( Fig. 388 View FIGURES 384 – 389 ). as1 present. Basal apodeme largely rectangular, widening slightly distally, with rounded anterior end indented medianly. Proximal mesosome trapezoidal, narrow anterior, overlapping basal apodeme. Mesosomal lobes wide, rounded, with distal section elongated medianly to approach distal parts of parameres. Thickened transversal nodi at about half length of lobed; ames and pmes of roughly equal length, forming distally converging rows. Parameral heads finger-like. Parameral blades broad, tapering only distally.
Sturnidoecus mon species-group ( Fig. 389 View FIGURES 384 – 389 ). as1 absent. Basal apodeme rectangular, anterior end rounded. Proximal mesosome square-shaped, anterior margin irregular, overlapping basal apodeme. Gonopore as crescentshaped thickening at terminal margin of mesosome; anterior margin of gonopore serrated. Mesosomal lobes rectangular, squat; 2 ames much longer than 3 pmes, the latter sensilla. ames submedian, near anterior end of mesosomal lobes; pmes submedian, lateral to gonopore. Parameral heads rectangular. Parameral blades broad, tapering only distally, straight.
Sturnidoecus pastoris species-group ( Fig. 390 View FIGURES 390 – 395 ). as1 present. Basal apodeme truncated ovoid, anterior margin rounded. Proximal mesosome as in Fig. 390 View FIGURES 390 – 395 , wider anteriorly, with uneven anterior margin, overlapping basal apodeme. Mesosomal lobes oval, slender, with convoluted thickenings and large hooks on ventral side; 2 sublateral ames much longer than 3 submedian pmes l setae together form distally converging rows. Parameral heads roughly triangular. Parameral blades gently tapering.
Sturnidoecus porphyrogenitus species-group ( Fig. 391 View FIGURES 390 – 395 ). as1 present. Basal apodeme broadly oval, wider proximally, anterior margin rounded. Proximal mesosome trapezoidal, wider anteriorly, overlapping basal apodeme. Mesosomal lobes wide, roughly rectangular, with sinuous lateral margins; 2 ames and 3 pmes of roughly equal length; ames lateral to pmes. Parameral heads roughly rectangular. Parameral blades slender, angular.
Sturnidoecus senegalensis species-group ( Figs 392–393 View FIGURES 390 – 395 ). as1 present. Basal apodeme roughly rectangular, anterior end rounded. Proximal mesosome triangular, wider anteriorly with thickened anterior margin, not overlapping basal apodeme. Mesosomal lobes slender, elongated, nearly as long as parameres. One or two hookshaped bulges on ventral side ( Fig. 393 View FIGURES 390 – 395 ); 4 ames shorter than single pmes; sets form distally converging rows much anterior to gonopore. Parameral heads bent and blunt. Parameral blades elongated, straight, tapering only distally.
Sturnidoecus simpl ex species-group ( Fig. 394 View FIGURES 390 – 395 ). as1 present. Basal apodeme rectangular, anterior margin rounded. Proximal mesosome roughly quadratic, overlapping basal apodeme. Mesosomal lobes roundedly rectangular, with wide thickenings laterally and distally; 2 sublateral ames much longer than 3 submedian pmes, and sets widely separated. Parameral heads rounded, with thumb-like extension on anterior end. Parameral blades lobe-like, short.
Sturnidoecus somnodraco species-group ( Fig. 395 View FIGURES 390 – 395 ). as1 absent. Basal apodeme rectangular, anterior margin rounded. Proximal mesosome as in Fig. 395 View FIGURES 390 – 395 , overlapping basal apodeme; lateral margins with prominent point at half length. Mesosomal lobes slender, with large thickenings laterally; 2 ames sublaterally on each side. No pmes visible. Parameral heads bluntly quadratic. parameral blades triangular, folded medianly; lateral margin with prominent point; parameres short.
Sturnidoecus sturni species-group ( Fig. 396 View FIGURES 396 – 398 ). as1 present. Basal apodeme truncated oval, anterior margin rounded. Transverse band along distal margin of basal apodeme. Proximal mesosome bilobed, overlapping basal apodeme. Mesosomal lobes slender, somewhat elongated, with large hooks distally; 2 sublateral ames much longer than 3 submedian pmes, which are often sensilliform and hard to see; ames and pmes much separated. Parameral heads fingerlike, slender. Parameral blades broad, straight, tapering only distally.
Sturnidoecus wittei species-group ( Fig. 397–398 View FIGURES 396 – 398 ). as1 present. Basal apodeme almost quadratic, anterior margin flat. Proximal mesosome widely trapezoidal with thickened anterior margin, overlapping basal apodeme. Mesosomal lobes slender, elongate, approaching distal parameres. Elongated section fused medianly; 4 ames much longer than single pmes, sets much separated but forming distally converging rows. Prominent ventral horns on anterior third of mesosome ( Fig. 398 View FIGURES 396 – 398 ). Parameral heads broad and rounded. Parameral blades long, broad, tapering only near distal tips.
Host distribution. Sturnidoecus is widely distributed on members of several families of birds, primarily Sturnidae , Turdidae , Ploceidae , and Malaconotidae , but some species occur on several other host families (see Appendices I and II), and we have seen undescribed material from additional host families (unpub. data) including Campephagidae , Estrildidae , Icteridae , Lybiidae , Mimidae , Motacillidae , Nectariniidae , Stenostiridae , Timaliidae , Thraupidae . The distribution in the New World appears entirely limited to thrushes and introduced host species (such as Sturnus vulgaris ), but in Sub-Saharan Africa Sturnidoecus is widely distributed on several host families. Sturnidoecus appears to be replaced by Schizosairhynchus on several South-East Asian sturnid genera, such as Aplonis and Mino [the “South-East Asian/Pacific Starlings ” of Lovette & Rubenstein (2007)]. Intriguingly, Sturnidoecus has not been found on either Mimidae or Rhabdornithidae , both of which are closely related to the Sturnidae and Buphagidae ( Zuccon et al. 2006; Lovette & Rubenstein 2007).
Geographical distribution. Near global, but in some areas primarily on introduced hosts.
Included species
(arranged by species group)
Sturnidoecus australafricanus group
* Sturnidoecus australafricanus new species
Sturnidoecus basilewskyi group
* Sturnidoecus basilewskyi Tendeiro, 1963: 15 [1]
Sturnidoecus basilewskyi minor Tendeiro, 1963: 17 new synonymy Sturnidoecus lopesi Tendeiro, 1963: 21 new synonymy
* Sturnidoecus galbula Tendeiro, 1963: 29
Sturnidoecus neointermedius Price, Hellenthal & Palma, 2003: 243 Sturnidoecus intermedius Tendeiro, 1963: 37 nec Sturnidoecus intermedius Ansari, 1955a
* Sturnidoecus sexualis Tendeiro, 1963: 34
* Sturnidoecus textoris Tendeiro, 1964: 196
* Sturnidoecus xanthops Tendeiro, 1963: 43
Sturnidoecus clayae group
* Sturnidoecus avivorax Ansari, 1968: 5 * Sturnidoecus clayae Ansari, 1968: 5
Sturnidoecus meinertzhageni group
* Sturnidoecus meinertzhageni Ansari, 1968: 5
Sturnidoecus mon group
* Sturnidoecus mon new species










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Sturnidoecus Eichler, 1944
| Bush, Sarah E. 2017 |
Sturnidoecus neointermedius
| Price 2003: 243 |
| Tendeiro 1963: 37 |
Sturnidoecus avivorax
| Ansari 1968: 5 |
| Ansari 1968: 5 |
Sturnidoecus meinertzhageni
| Ansari 1968: 5 |
Sturnidoecus textoris
| Tendeiro 1964: 196 |
Sturnidoecus basilewskyi
| Tendeiro 1963: 15 |
Sturnidoecus basilewskyi minor
| Tendeiro 1963: 17 |
| Tendeiro 1963: 21 |
Sturnidoecus galbula
| Tendeiro 1963: 29 |
Sturnidoecus sexualis
| Tendeiro 1963: 34 |
Sturnidoecus xanthops
| Tendeiro 1963: 43 |
Turdinirmus
| Eichler 1951: 41 |
Penenirmus
| Eichler 1944: 81 |
| Clay 1938: 73 |
Philopterus
| Nitzsch 1818: 288 |
| Nitzsch 1818: 289 |
| Nitzsch 1818: 291 |