
Nihonella, Ballarin & Yamasaki, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.733.1215 |
|
publication LSID |
lsid:zoobank.org:pub:3675D40A-5DE0-49CE-9795-F4FCF13B17CB |
|
DOI |
https://doi.org/10.5281/zenodo.4472795 |
|
persistent identifier |
https://treatment.plazi.org/id/05F43F8A-6FBC-4A95-98FF-56CEFACF73DB |
|
taxon LSID |
lsid:zoobank.org:act:05F43F8A-6FBC-4A95-98FF-56CEFACF73DB |
|
treatment provided by |
Plazi |
|
scientific name |
Nihonella |
| status |
gen. nov. |
Nihonella View in CoL View at ENA gen. nov.
urn:lsid:zoobank.org:act:05F43F8A-6FBC-4A95-98FF-56CEFACF73DB
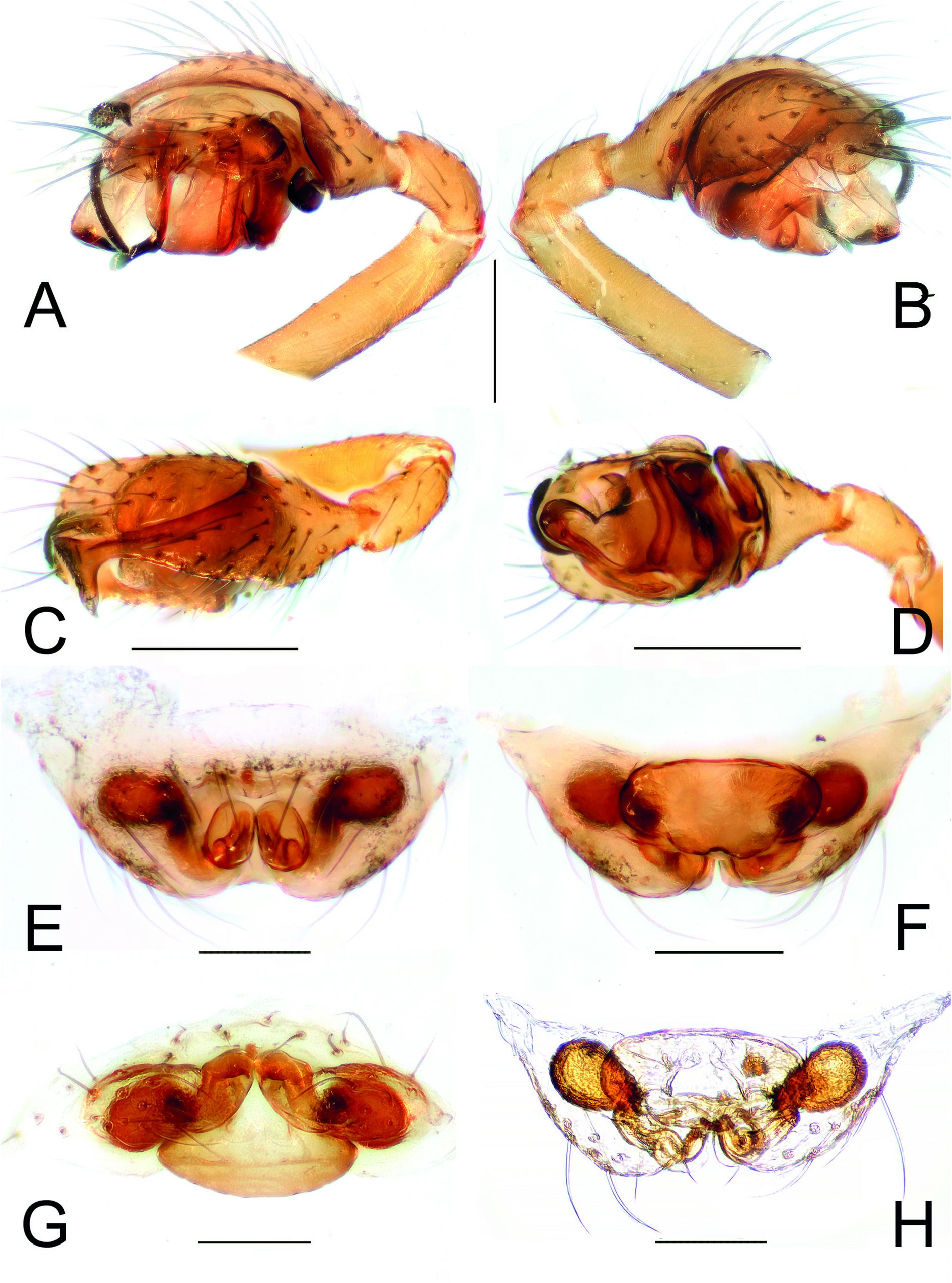
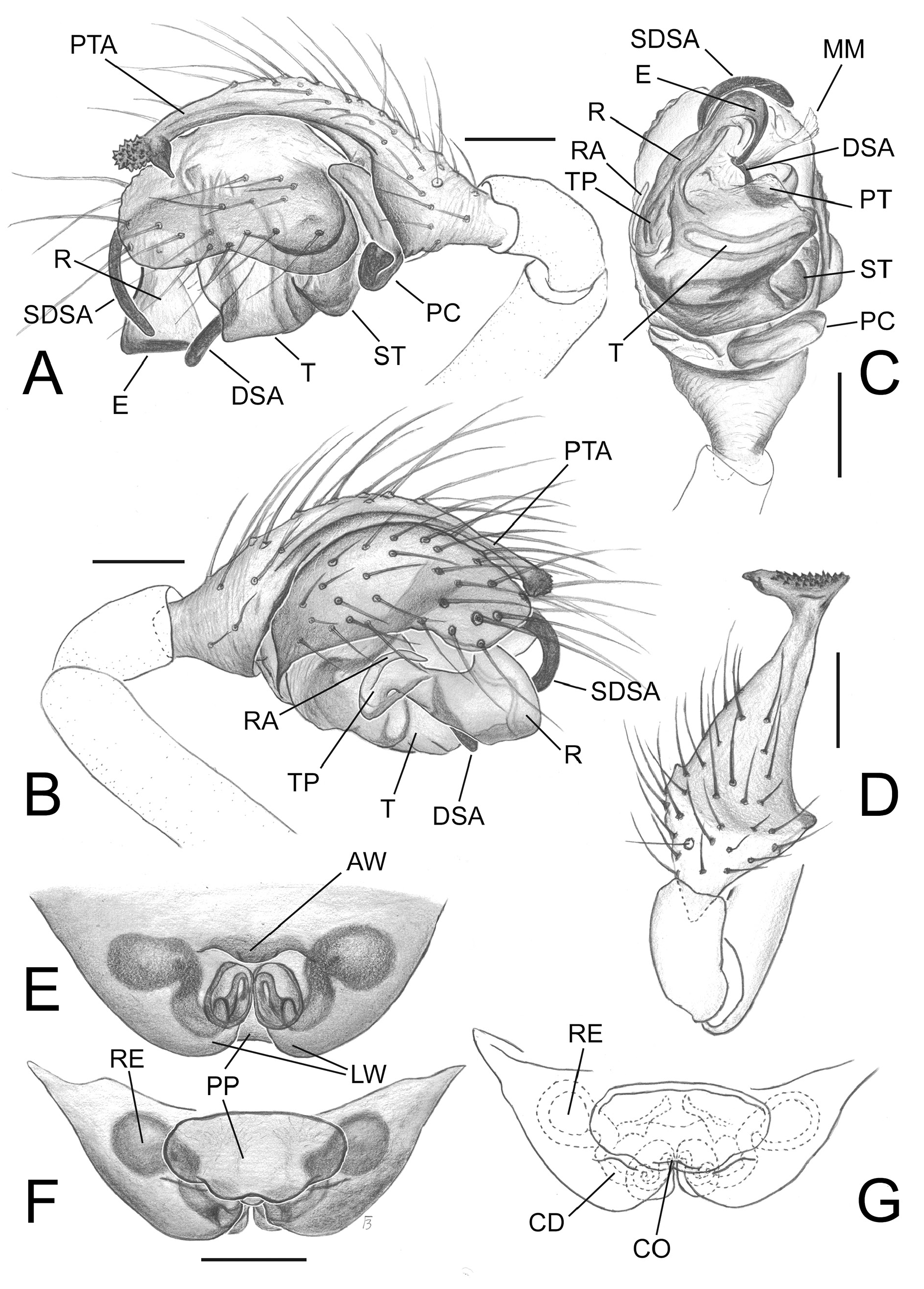
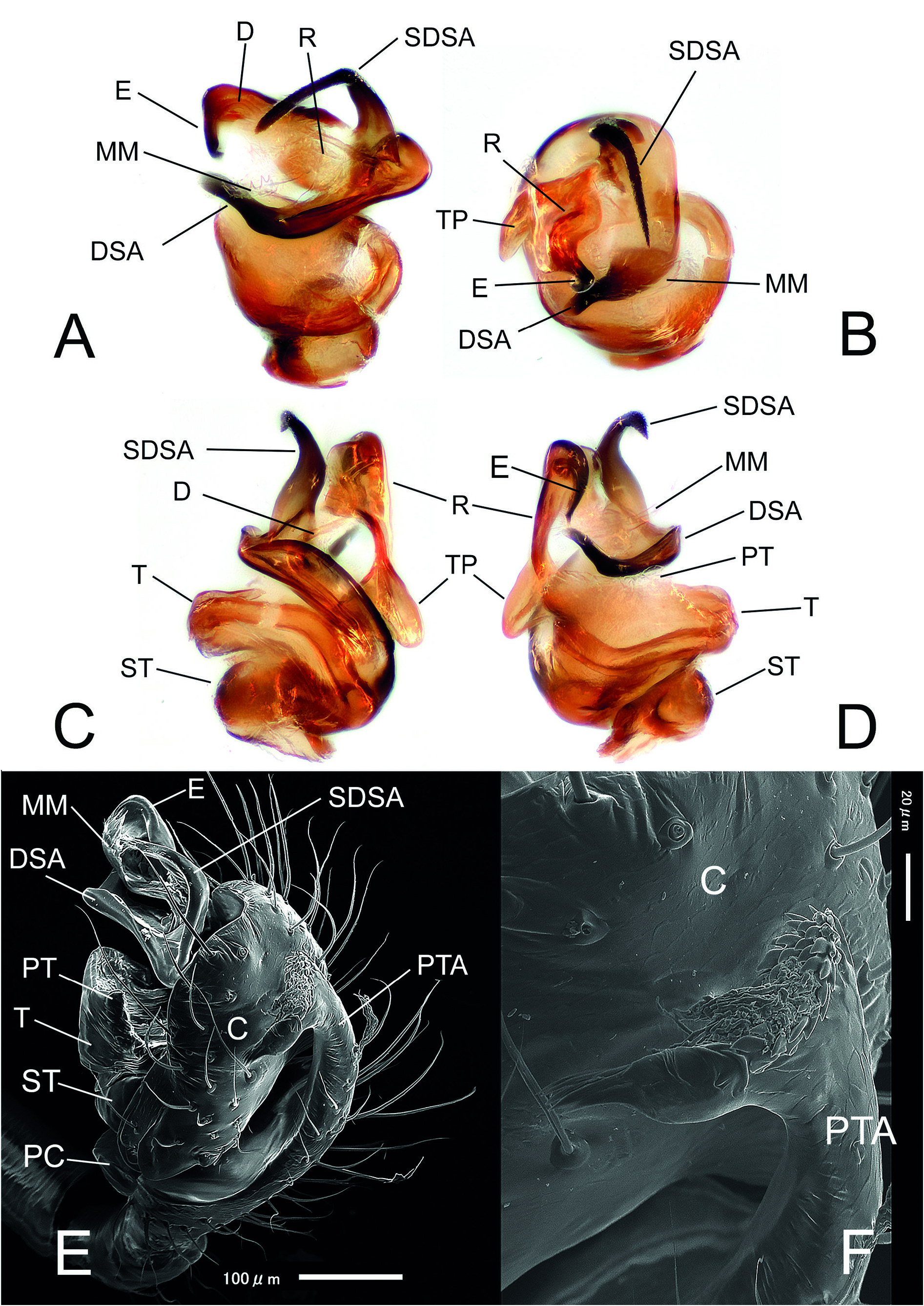
Figs 1 View Fig A–G, 2–4 View Fig View Fig View Fig ; Table 2 View Table 2
Type species
Nihonella View in CoL chika gen. et. sp. nov.
Diagnosis
The new genus is distinguished from any other genera belonging to the distal Erigoninae clade by the following unique combination of somatic and genitalic characters: Femur I with 1 prolateral spine; Tibia I with 1 dorsal spine; tibial spine formula: 1.1.1.1; male palp with a well-developed distal suprategular apophysis and a hypertrophic ʻsecondaryʼ DSA ( Figs 2 View Fig A–B, 3A–C, 4A–E) (usually presented in Erigoninae as a simple tooth and protruding form a different side of the DSA); a well-developed and uniquely-shaped prolateral tibial apophysis, the same length as the cymbium and partially covering it ( Figs 2 View Fig A–C, 3A–B, D, 4E–F). The unusual chaetotaxy and unique shape of the epigyne, with anteriorly converging lateral walls and two distinct, flat, ovoid inflations of the copulatory ducts, also distinguishes the female of this genus from females of any other genera in distal Erigoninae .
Etymology
The generic name is a combination of the word ʻ Nihon ʼ and the Latin suffix ʻ -ella ʼ. The former refers to the country of Japan where the genus is endemic; the latter is the feminine suffix of ʻ -ellus ʼ commonly used in Latin to form the feminine diminutive of a noun. Name in apposition, feminine in gender.
Species included
Only the type species Nihonella chika gen. et. sp. nov.
Taxonomic remarks
The morphology of Nihonella gen. nov. suggests it may be closely related to the species of the Savignia group. However, the presence of numerous differences in genitalia shape and somatic characters does not allow us to include the new genus within this group of species. A close but distinct relationship with the Savignia group is also supported by the molecular analysis (see Fig. 5 View Fig ).
Nihonella gen. nov. male palps have a general morphology similar to those found in some genera of the Savignia group (e.g., Araeoncus Simon, 1884 or Diplocephalus Bertkau, 1883 ). They share a similar shape in the embolic division: a long, modified palpal tibia, and a well-developed DSA. However, the new genus shows a distinct hypertrophy of the SDSA, which is extremely long and clearly protruding outside the frontal part of the palpal bulb ( Figs 2 View Fig A–B, D, 3A–C, 4A–E). Within the subfamily Erigoninae , some genera belonging to the Savignia group (sensu Millidge 1977) have a large DSA and a tooth-like SDSA (e.g., Alioranus Simon, 1926 , Dactylopisthes Simon, 1884 , Delorrhipis Simon 1885 , Savignia Blackwall, 1833 , etc., see Millidge 1977: figs 128, 135–136, 139). However, none of the Savignia group display a SDSA as strongly developed as in Nihonella gen. nov. Females of Nihonella gen. nov. have an epigyne with two anteriorly converging lateral walls, which resembles the female genitalia, of most of genera within the Savignia group (e.g., Araeoncus Simon, 1884 , Diplocephalus Bertkau, 1883 , Erigonella Dahl, 1901 , Savignia Blackwall, 1833 , etc.). Nevertheless, both males and females of Nihonella gen. nov. have a highly distinctive chaetotaxy which strongly differ from the chaetotaxy usually found in species belonging to the Savignia group (1.1.1.1 vs 2.2.1.1). Although some species included in this group may occasionally have a tibial spine formula of 1.1.1.1, this usually only occurs in males and as a consequence of the reduction of the distal spines in tibia I and II (e.g., Araeoncus crassipes Heimer & Nentwig, 1991 = 1.1.1.1, A. humilis (Blackwall, 1841) = 0.0.1.1: Tanasevitch, in litteris). An exception is the genus Microctenonyx Dahl, 1886 , the female of which has a tibial spine formula of 1.1.1.1. However, Microctenonyx can be easily distinguished from Nihonella gen. nov. by the large genetic distance between the two genera (see Table 2 View Table 2 ), and by the shape of the epigyne and male palp (short SDSA, different shape of epigyne and internal ducts, see Figs 2 View Fig A–H, 3A–D vs Millidge 1977: fig. 140 and Bosmans 2007: figs 111–115).
Distribution
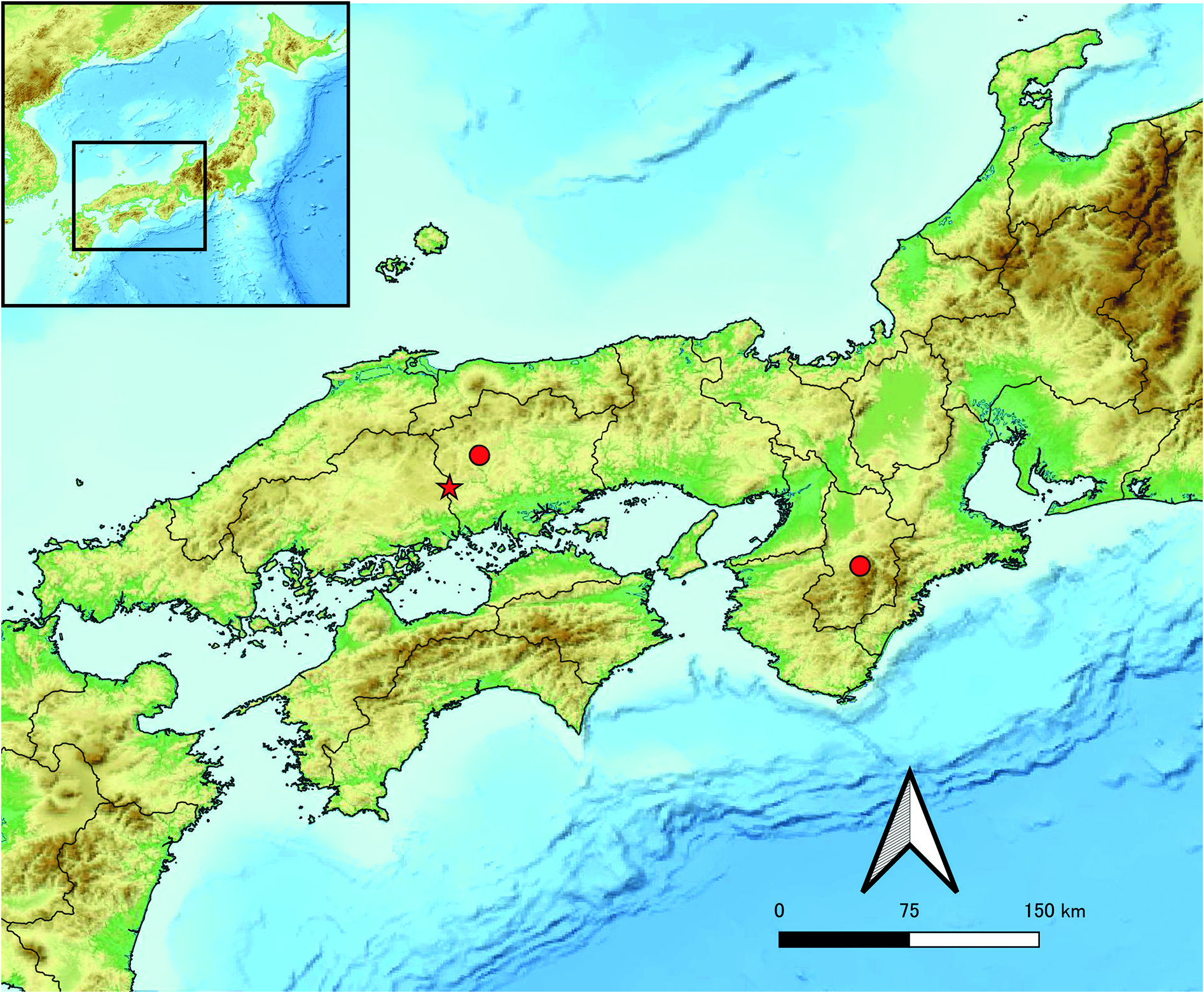
Endemic to Western Honshu, Japan. Currently known from three caves only ( Figs 1H View Fig , 6 View Fig ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Erigoninae |