
Eulepetopsis crystallina, Chen & Zhou & Watanabe & Zhang & Wang, 2022
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlab081 |
|
publication LSID |
lsid:zoobank.org:pub:6334CD15-D490-496F-802A-F162B4FF8A21 |
|
DOI |
https://doi.org/10.5281/zenodo.5800010 |
|
persistent identifier |
https://treatment.plazi.org/id/88425156-653A-FFC4-FC49-A2660881F951 |
|
treatment provided by |
Plazi |
|
scientific name |
Eulepetopsis crystallina |
| status |
sp. nov. |
EULEPETOPSIS CRYSTALLINA View in CoL SP. NOV.
( FIGS 7–9 View Figure 7 View Figure 8 View Figure 9 )
ZooBank registration: urn:lsid:zoobank.org:act:
Neolepetopsidae View in CoL gen. sp. – Hashimoto et al., 2001: 720, table 1.
Eulepetopsis View in CoL – Van Dover et al., 2001: 821, table 2.
Eulepetopsis sp. – Watanabe & Beedessee, 2015: 207, table 16.1; Sun et al., 2020: 8, table 1; Kim et al., 2020: supplementary table 1.
Eulepetopsis sp. ‘SWIR’ – Zhou et al., 2018: 7, table 1.
‘An unnamed species known from the Kairei Vent Field’ – Warén et al., 2006: 83.
Diagnosis: Typical-sized Eulepetopsis ≤ 14.0 mm SL. Radula with sturdy pluricuspid teeth carrying a clear lateral projection mid-shaft, in addition to a finely serrated cutting edge.
Type locality: On active vent chimney, Kairei vent field, Central Indian Ridge, 25°19.2315′S, 70°2.4187′E, 2424 m deep GoogleMaps .
Type material: Holotype (NSMT-Mo 79222; Fig. 7A View Figure 7 ) fixed in 10% buffered formalin and stored in 70% ethanol, SL 8.4 mm, SW 6.0 mm, Kairei vent field, Central Indian Ridge, 25°19.2315′S, 70°2.4187′E, 2424 m deep, collected by suction sampler, R / V Yokosuka cruise YK16-E02, DSV Shinkai 6500 dive #1449, 13 February 2016 GoogleMaps . Paratype 1 (NSMT-Mo 79223; Fig. 7B View Figure 7 ), fixed in 10% buffered formalin and stored in 70% ethanol, SL 8.7 mm, SW 8.0 mm. Paratype 2 (NSMT-Mo 79224), 99% ethanol, used for DNA extraction and sequencing, SL 10.8 mm, SW 8.0 mm. Paratype 3 (NSMT-Mo 79225; Fig. 7C View Figure 7 ), fixed in 10% buffered formalin and stored in 70% ethanol, SL 8.3 mm, SW 5.8 mm. All paratypes are from the same lot as the holotype.
Materials examined: One specimen (NSMT-Mo 79226; Fig. 7D View Figure 7 ), Edmond vent field, Central Indian Ridge, 99% ethanol, SL 11.8 mm, SW 8.6 mm, 23°52.6621′S, 69°35.7959′E, 3279 m deep, collected by suction sampler, R / V Yokosuka cruise YK16-E02, DSV Shinkai 6500 dive #1457, 26 February 2016, covered in thick sulfide layer, now removed, in part, to reveal shell surface. Six specimens ( RSIO 35734 ; Fig. 7E View Figure 7 ), GoogleMaps Tiancheng vent field, Southwest Indian Ridge, SL 11.2–14.1 mm, SW 8.2–11.0 mm, 63°55.398′E, 27°51.030′S, 2682 m deep, collected by a seven-function manipulator, R / V Xiangyanghong 9 cruise DY35, HOV Jiaolong dive 87, 23 December 2014 GoogleMaps . Three specimens ( RSIO 38215 ; Fig. 7F View Figure 7 ), Wocan vent field, Carlsberg Ridge, SL 10.5–14.0 mm, SW 7.5–10.2 mm, covered in thin layer of sulfide deposits, 60°31.8′E, 6°21.6′N, 2920 m deep, collected by a seven-function manipulator, R / V Xiangyanghong 9 cruise DY38, HOV Jiaolong dive 129, 14 March 2017 GoogleMaps .
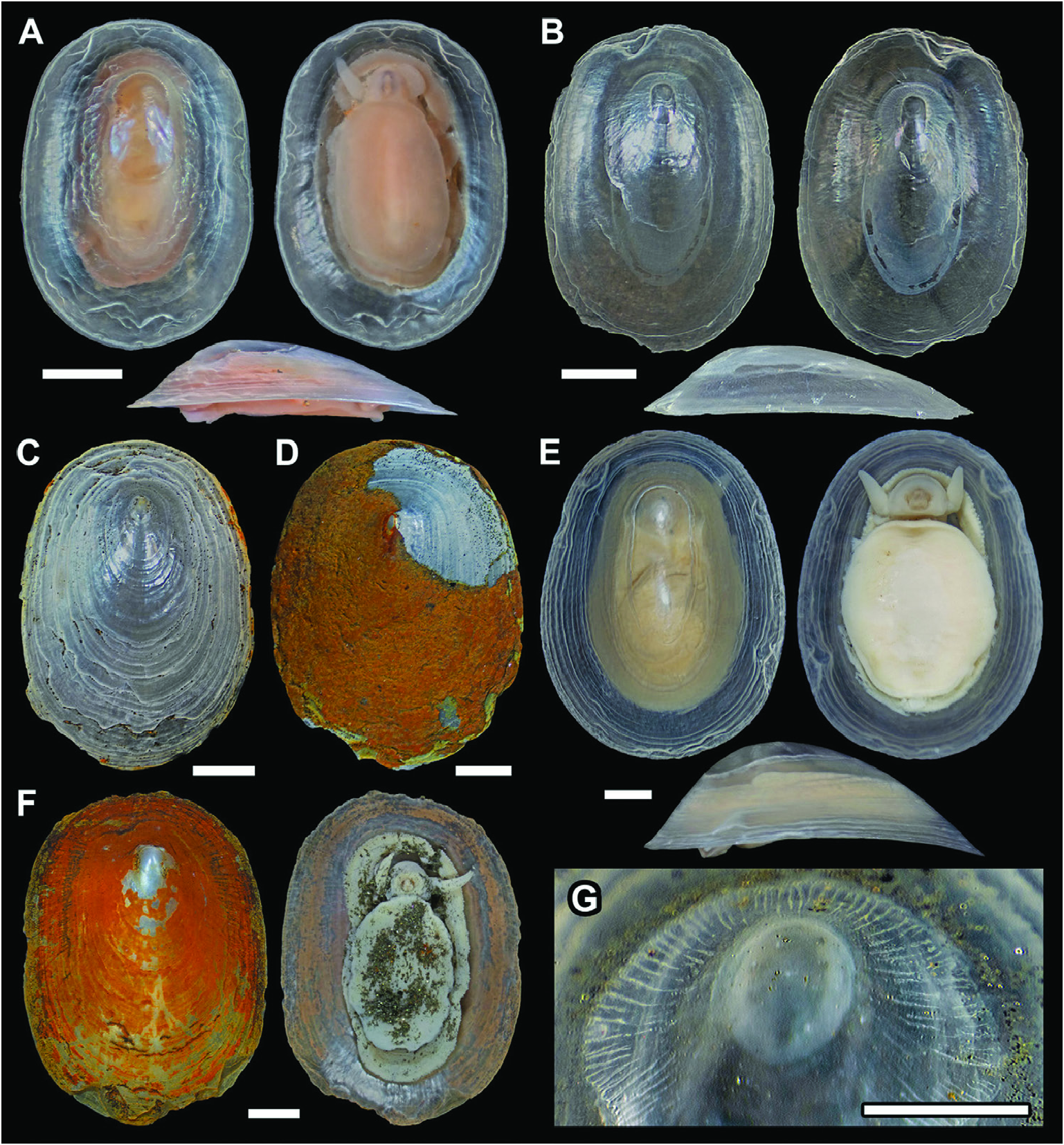
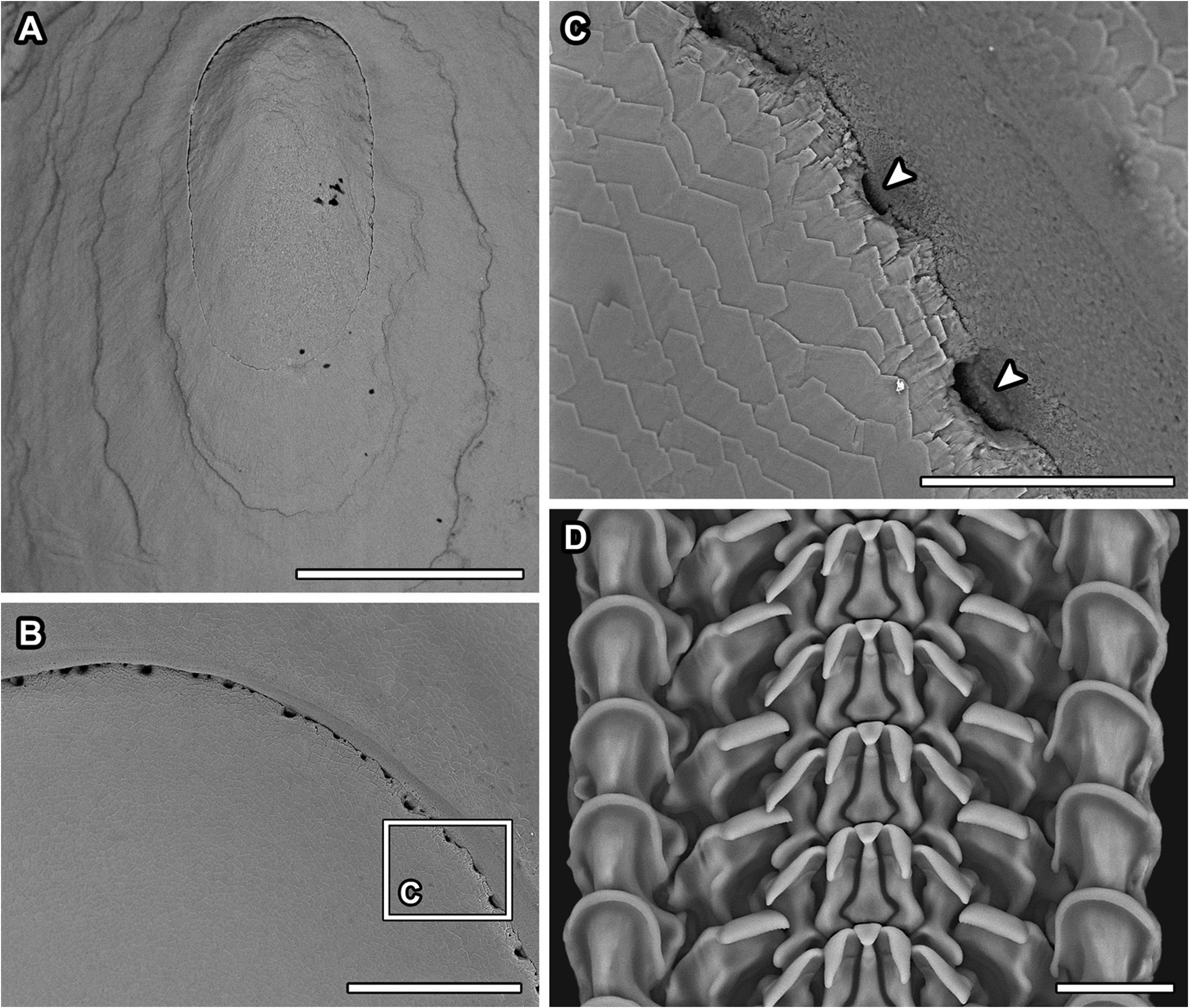
Description: Shell ( Fig. 7 View Figure 7 ) thin, fully transparent, with thin layer of periostracum where not corroded. Shell length oblong oval; slightly narrower at anterior end than posterior end, more so in larger specimens ( Fig. 7E View Figure 7 ). Maximum known shell sizes at SL 14.0 mm, SW 10.2 mm. Shell profile low, flat, with margin almost aligned along one plane in smaller specimens, becoming more uneven in larger ones ( Fig.7 View Figure 7 ). Protoconch unknown; inner surface of protoconch sealed in specimens with lost protoconchs. Apex situated on midline anteriorly, about one-quarter of shell length from anterior edge. Shell surface almost completely smooth except for concentric growth lines ( Fig. 7C View Figure 7 ); area near apex corroded, with uneven lines ( Fig. 8A View Figure 8 ). Inner surface of shell showing muscle scars ( Fig. 7B View Figure 7 ); area near apex surrounded by a series of pores going into interior of shell towards apex ( Fig. 8B, C View Figure 8 ), seen as long radial streaks by transparency under optical microscopy ( Fig. 7G View Figure 7 ). Zigzagged crystal edges visible on inner surface of shell with electron scanning microscopy ( Fig. 8C View Figure 8 ).
Radula ( Fig. 8D View Figure 8 ) with sturdy rachidian and two laterals, pluricuspid tooth, two marginals on either side. Cusps not well mineralized. Rachidian well supported, with laterally expanded base; shaft of moderate length, slowly tapering apically, ending in narrow, triangular, overhanging cusp. Inner laterals with elongate, tapering triangular cusps on a solid shaft carrying strong indentation to accommodate lateral supports of rachidian tooth. Outer laterals about twice as broad as inner laterals; shafts with weak indentation to accommodate inner laterals, each carrying one prominent lateral projection near base. Pluricuspid robust, more than twice as wide as outer laterals, with prominent lateral, mid-shaft projection. Overhanging cusp of pluricuspid broad, with numerous fine serrations decreasing in strength outward. Laterals and pluricuspid decrease in cusp positions outward from the rachidian. Inner marginal well formed, as broad as pluricuspid, with narrow, smooth, semicircular, overhanging cutting edge. Second marginal vestigial, found slightly outside of inner marginal.
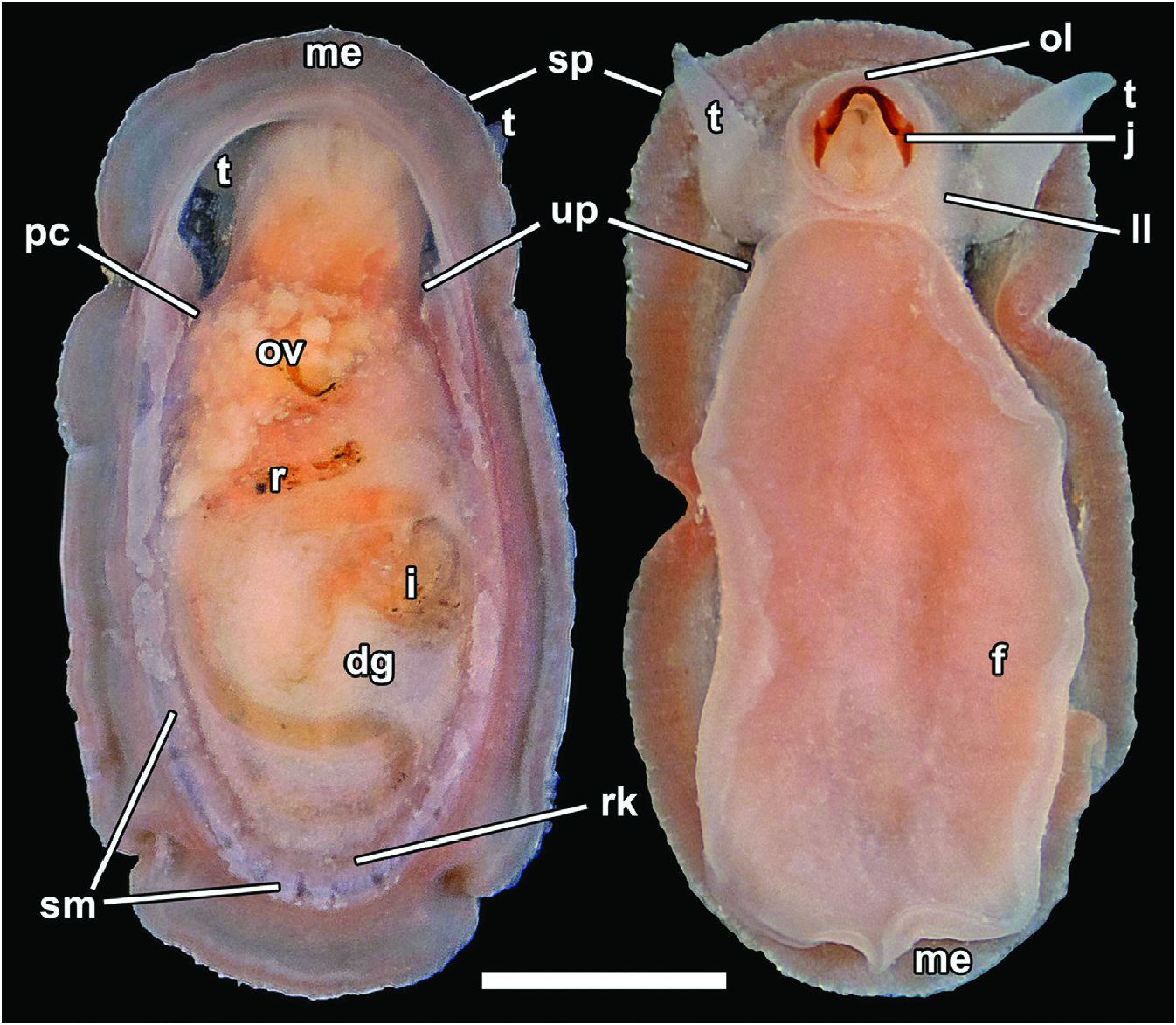
Soft parts are shown in Figure 9 View Figure 9 . Cephalic tentacles simple conical, without appendages, elongate, tapered. No external evidence for eyes. Oral disc with muscular outer lip, surrounded by moderately developed labial lobe. Well-developed, dorsally arched jaw present, often seen projecting from mouth in preserved specimens. Sole of foot oval, large, with unciliated rim demarcated by deep groove from sole. Epipodium lacking. Shell muscle U-shaped, separated into numerous oblong muscle bundles along the posterior three-quarters of body, with length of bundles decreasing posteriorly. Mantle edge with numerous fine, presumably sensory papillae. Mantle cavity shallow, extending to slightly shy of one-third of body length. Heart monotocardian, with auricle anterior of ventricle (seen by transparency), located within pericardium on left mantle roof. Ctenidium lacking. Sexes separate; gonad located ventrally along mid-body, partly visible from dorsal view slightly posterior to pericardium. Left kidney minute; sizeable right kidney positioned at posterior of body. Urogenital papillae on right mantle roof on right side of anus. Intestine much wider anteriorly, looping twice before emerging at posterior end of body as rectum. Rectum runs towards anterior left before turning to anterior right and finally emerges on right mantle roof. Intestine and stomach entirely embedded within voluminous digestive gland, comprising numerous tubular structures. Operculum lacking.
Etymology: From Latin crystallinum, crystal-like, named for its highly transparent shell.
Distribution: Known from a number of hydrothermal vent fields across Carlsberg Ridge (Wocan field), Central Indian Ridge (Kairei and Edmond fields) and Southwest Indian Ridge (Tiancheng field). Given its distribution range from the examined materials, Eulepetopsis recorded at both Solitaire ( Nakamura et al., 2012; Watanabe & Beedessee, 2015) and Onnuri ( Kim et al., 2020) fields are most likely additional records of this species.
Remarks: Eulepetopsis crystallina is similar morphologically to E. vitrea , the only other known species in the genus, described from vents on the East Pacific Rise. Both have highly transparent shells and similar anatomical features ( Fretter, 1990), but are separable based on radula morphology, most notably the pluricuspid teeth. In E. crystallina , the pluricuspid has a prominent mid-shaft projection, which is lacking in that of E. vitrea ( McLean, 1990; Warén & Bouchet, 2001). The finely serrated cutting edge is another feature not mentioned in E. vitrea ; the pluricuspid teeth as a whole are much stronger and broader at the base in E. crystallina than in E. vitrea ( McLean, 1990; Warén & Bouchet, 2001). Shafts of all teeth in E. crystallina are also noticeably longer than in E. vitrea ( Warén & Bouchet, 2001) .
GENETIC SUPPORT
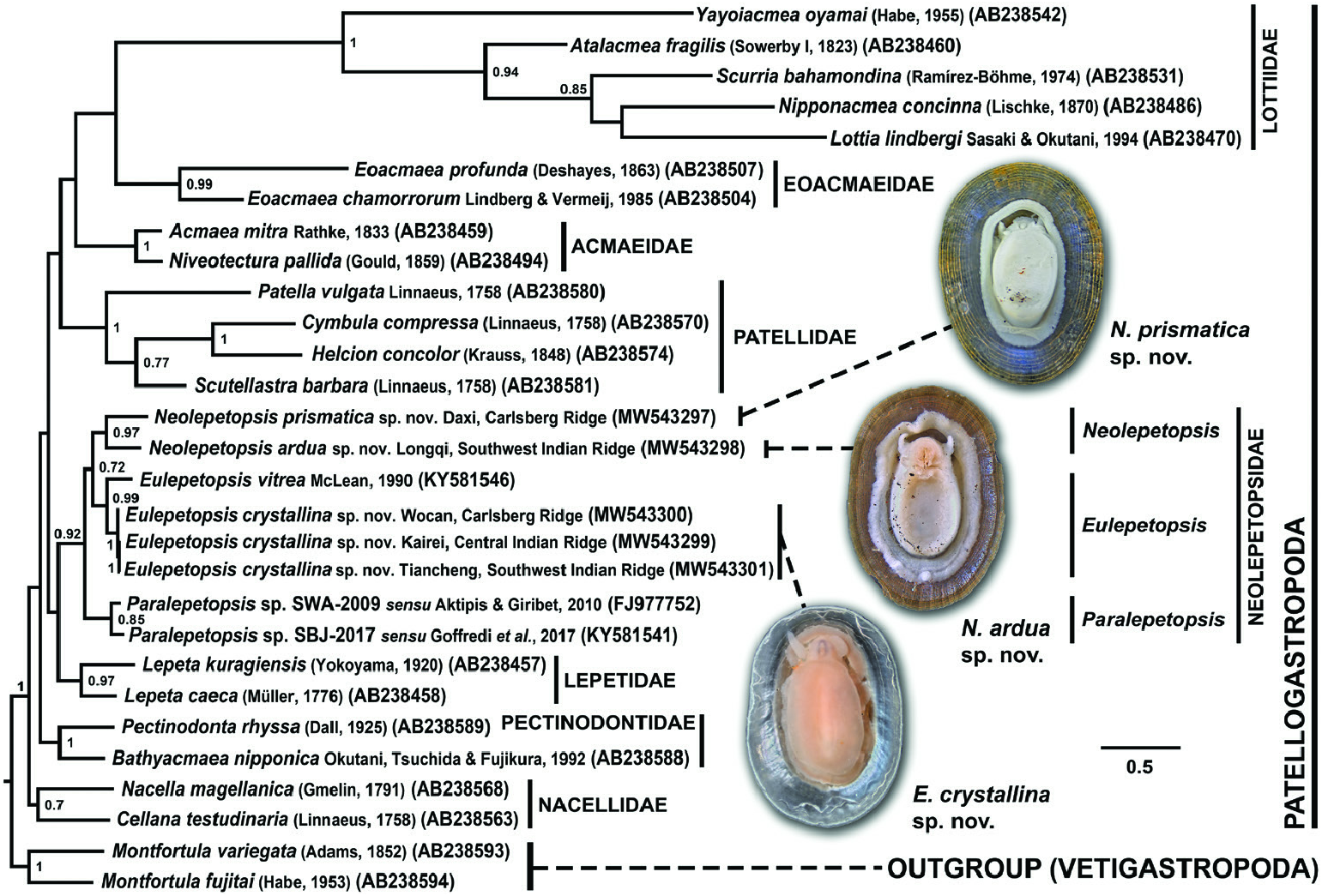
The consensus tree for Patellogastropoda from phylogenetic reconstruction by Bayesian inference using first and second codon positions of a 472 bp alignment in the barcoding region is shown in Figure 10 View Figure 10 . Neolepetopsis prismatica was recovered as sister to N. ardua with strong support [Bayesian posterior probability (BPP) = 0.97], with the clade interpreted as genus Neolepetopsis . The three sequences of E. crystallina included, one from each mid-ocean ridge, were recovered as a fully supported clade corresponding to the new species. This was sister to E. vitrea , with the two species forming a strongly supported clade (BPP = 0.99) representing genus Eulepetopsis . Eulepetospsis was recovered as sister to Neolepetopsis with moderate support (BPP = 0.72), with this pair in turn being sister to a moderately supported (BPP = 0.85) Paralepetopsis containing two (undescribed) species. This means that Paralepetopsis was recovered in a basal position within Neolepetopsidae , which was recovered as a well-supported (BPP = 0.92) clade containing the three abovementioned genera.
At the level of families within Patellogastropoda , all currently established patellogastropod families ( Nakano & Ozawa, 2007; Aktipis & Giribet, 2010; Nakano & Sasaki, 2011; Goffredi et al., 2017) were recovered as moderately to well-supported clades (BPP = 0.7–1.0), but all sister relationships between families were not well supported. Neolepetopsidae was recovered as sister to Lepetidae , but this relationship was not well supported (BPP = 0.52).
The genetic distances (K2P distances), estimated using 472 bp of the COI gene, among neolepetopsid taxa with suitable data available are shown in Table 1 View Table 1 . The genetic distance for the three specimens of E. crystallina included, one from each ridge system, was 0.21–0.85%. This is much lower than the distance between these and E. vitrea , which ranged between 9.79 and 10.30%. The genetic distance between N. prismatica and N. ardua was estimated at 5.54%, and that between the two undescribed Paralepetopsis species ( Aktipis & Giribet, 2010; Goffredi et al., 2017) was 12.26%. The average genetic distance between species assigned to the same genera was 9.59% (range 5.54–12.26%), and between species of different genera it was 13.95% (12.27–15.09%).
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Patellogastropoda |
|
Order |
|
|
Family |
|
|
Genus |
Eulepetopsis crystallina
| Chen, Chong, Zhou, Yadong, Watanabe, Hiromi Kayama, Zhang, Ruiyan & Wang, Chunsheng 2022 |
Eulepetopsis sp.
| Zhou Y & Zhang D & Zhang R & Liu Z & Tao C & Lu B & Sun D & Xu P & Lin R & Wang J & Wang C 2018: 7 |
Eulepetopsis sp.
| Sun J & Zhou Y & Chen C & Kwan YH & Sun Y & Wang X & Yang L & Zhang R & Wei T & Yang Y & Qu L & Sun C & Qian PY 2020: 8 |
| Watanabe H & Beedessee G 2015: 207 |
Neolepetopsidae
| Hashimoto J & Ohta S & Gamo T & Chiba H & Yamaguchi T & Tsuchida S & Okudaira T & Watabe H & Yamanaka T & Kitazawa M 2001: 720 |
Eulepetopsis
| Van Dover CL & Humphris SE & Fornari D & Cavanaugh CM & Collier R & Goffredi SK & Hashimoto J & Lilley MD & Reysenbach AL & Shank TM & Von Damm KL & Banta A & Gallant RM & Gotz D & Green D & Hall J & Harmer TL & Hurtado LA & Johnson P & McKiness ZP & Meredith C & Olson E & Pan IL & Turnipseed M & Won Y & Young CR 3 rd & Vrijenhoek RC 2001: 821 |