
Neoeubria inbionis, Shepard & Barr, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3811.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:C550F505-5BE1-422D-A196-1081B4CB6C04 |
|
DOI |
https://doi.org/10.5281/zenodo.5116331 |
|
persistent identifier |
https://treatment.plazi.org/id/8C3587E0-FF88-AF00-638F-AD8FFDC5A1AF |
|
treatment provided by |
Felipe |
|
scientific name |
Neoeubria inbionis |
| status |
sp. nov. |
Neoeubria inbionis sp. n.
Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 25–27 View FIGURE 25 View FIGURE 26 View FIGURE 27
Type material. Holotype (male): COSTA RICA: Guanacaste Prov., Parque Nacional Rincón de la Vieja, Las Pailas Trail , 14-VI-2001, William D. Shepard, leg. // reared from pupa collected on wood in seep basin // HOLOTYPE Neoeubria inbionis Shepard & Barr [red label]. Deposited in INBC . Allotype (female): locality data same as holotype // ALLOTYPE Neoeubria inbionis Shepard & Barr [red label]. Deposited in INBC . Paratypes (2 M & 5 F): ECUADOR: Napo Prov., Huahua Sumaco, Km 44 on Hollin-Loreto Rd. , XII-15-1989, Malaise Trap, MS/ JS Wasbauer, H. Real // CALIFORNIA STATE COLLN AGRICULTURE // PARATYPE Neoeubria inbionis Shepard & Barr [yellow label] (1 M) ( EMEC); data same, except XII-16-1989 (1 F) ( CSCA); data same, except XII-18-1989 (1 M) ( CSCA); data same, except XII-19-1989 (1 F) ( CSCA); data same, except XII-21-1989 (2 FF) ( CSCA, EMEC); data same, except XII-22-1989 (1 F) ( CSCA) .
Adult Description. Body oval; males ( Fig. 3 View FIGURE 3 ) smaller than females ( Figs. 1–2 View FIGURE 1 View FIGURE 2 , 4 View FIGURE 4 ); males 4.6–5.0 mm long and 2.75 mm wide, females 5.0– 5.6 mm long and 2.8–3.5 mm wide. Integument color medium brown, shiny where setae sparse; covered dorsally with a combination of different kinds of setae: widely-spaced, long, erect blond and dark brown setae; sparse, shorter, pale brown setae; and very dense, recumbent, pale blond setae forming a pattern of broad bands and large spots. Venter uniformly clothed in medium-length pale blond setae.
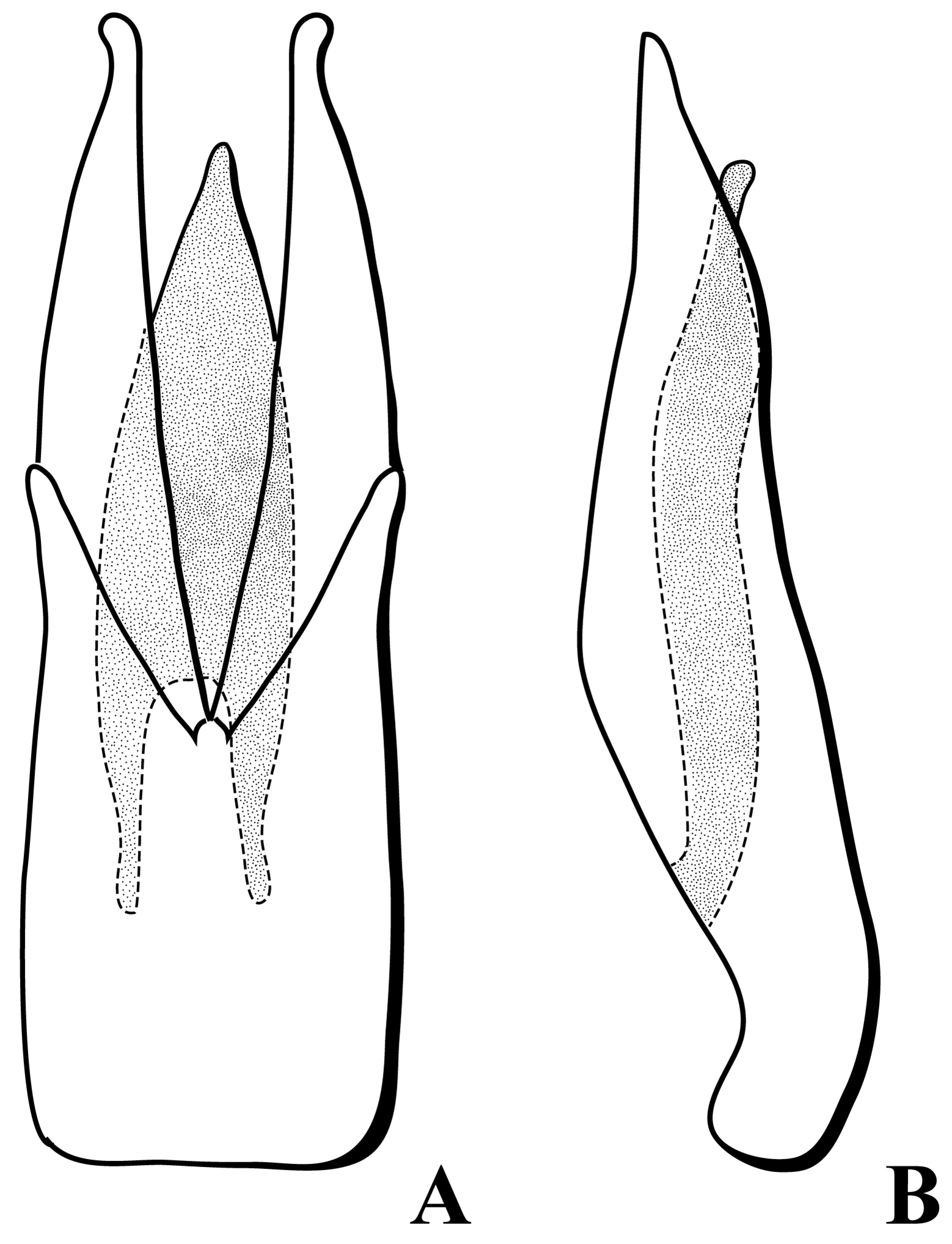
Aedeagus of trilobed type ( Fig. 25 View FIGURE 25 ) and lightly sclerotized. Basal piece long, reduced to ventral plate basally with lateral flanges that clasp the base of parameres. Parameres long, widest at apical three-fourths of aedeagus; tips narrow, curved laterally; dorsally conjoined just anterior to midlength. Penis lanceolate; shorter than parameres; tip slightly curved ventrally and laterally compressed; base deeply cleft.
Ovipositor ( Fig. 26 View FIGURE 26 ) with bacula long, 1.4 times as long as coxites, thin, gently curved; only partially sclerotized. Coxites 0.7 times as long as bacula; joined medially in basal half, divergent medially in apical half; laterally gently sinuate. Styli short, one-segmented. Long, thin accessory sclerite dorsally in basal third of membrane between coxites.
Immature specimens examined. COSTA RICA: Alajuela, Alta Masis, 9 VI 2000, Río San Lorenzo [WDS- A-1302] // William D. Shepard, leg. (1 larva) ; Guanacaste Prov., Parque Nacional Rincón de la Vieja, Las Pailas Trail , 18-I-2000, William D. Shepard & Cheryl B. Barr, collected on wood in seep basin [WDS-A-1283](24 larvae, 1 pupa) ; data same, except 14-VI-2001, William D. Shepard, leg. [WDS-A-1386] (3 larvae, 3 pupae); data same, except 15-VI-2003, William D. Shepard & Cheryl B. Barr [WDS-A-1541] (11 larvae); data same, except Quebrada Pailas below Catarata , 14-VI-2001, William D. Shepard, leg. [WDS-A-1387] (1 larva) . NICARAGUA: Río San Juan, Refugio Bartola, 10 VIII 2002, riffle 3, Río Bartola , William D. Shepard, leg. [WDS-A-1492] (1 larva) . PANAMA: Chiriquí, Fortuna Forest Res. , March 2004, Checo Colón-Gaud, leg. (1 larva) . All immature specimens are deposited in EMEC.
Etymology. Named in honor of INBio, the Instituto Nacional de Biodiversidad in Costa Rica. The case is genitive.
Distribution. Nicaragua, Costa Rica, Panama and Ecuador, based on adult and larval specimens.
Habitat. The type locality in Parque Nacional Rincón de la Vieja at an 780 m is a series of seeps in a small basin connected by a spring run to a narrow, slow-flowing forest stream which is a tributary of the Río Colorado. The entire area around the seeps and both streams is heavily forested and generally heavily shaded. In the seep basin the water is only about 2–3 cm deep over a substrate composed of a thick deposit of silt and fine detritus on which lie sticks and larger pieces of rotting, waterlogged wood ( Fig. 27 View FIGURE 27 ). In the basin the water is extremely slowmoving but in a couple of meters it begins to flow downhill in a narrow spring run which is crossed by the Las Pailas Trail between Stops 3 and 4.
Larvae and pupae of N. inbionis were collected on pieces of decomposing wood found in the seep basin. Larvae were positioned below the water’s surface and pupae were above. The water is likely hypoxic because of the fine organic detritus substrate, coupled with the lack of sunlight for aquatic photosynthesizers due to the heavily-shaded nature of the site. Possession of a plastron facilitates larval survival in this water. Neoeubria was the only psephenid present in the seep area, and the only other co-occurring aquatic byrrhoid Coleoptera was an unidentified ptilodactylid larva. Other arthropods present in the seep area included aquatic Hemiptera , Belostoma (Belostomatidae) and Ambrysus (Naucoridae) , and the crustacean Hyallela (Amphipoda) . No specimens were collected from the spring run formed by the seeps. A single larva was collected in a second, larger stream, Quebrada Pailas, a tributary of the Río Colorado, which is also located along the Las Pailas Trail.
The other sites at which larvae were collected by the senior author are also forest streams, although with rocky substrates and faster flow. Although the particular microhabitat of the larvae at these sites is unknown, at all of them submerged wood was common. Neoeubria inbionis has been collected at elevations ranging from as low as 40+ m in Nicaragua, to as high as 780 m in Costa Rica.
The Ecuadorian adults were all taken in Malaise traps which were set in a forested area to catch flies. Although we could obtain no further information beyond the label data, collection of adults via Malaise traps indicates that N. inbionis adults behave like other eubriine adults and fly near the aquatic habitat in which the larvae occur.
Phylogeny. In the recent phylogeny of the Psephenidae by Lee et al. (2007), Neoeubria is included as “Genus A.” In the most parsimonious tree Neoeubria is placed in a basal trichotomy within the subfamily Eubriinae . The trichotomy positions Neoeubria in one branch, Sclerocyphon + Tychepsephus in another branch, and the remainder of the eubriine genera in a third branch.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |