
Rhynchelmis aleutensis, Fend, 2005
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1093.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:AAFC9875-2AC9-4B69-AA2D-AEA06DB082EC |
|
persistent identifier |
https://treatment.plazi.org/id/8D2487CD-FFA6-FFCF-FF7A-FB092AFDF9D9 |
|
treatment provided by |
Felipe |
|
scientific name |
Rhynchelmis aleutensis |
| status |
sp. nov. |
Rhynchelmis aleutensis View in CoL n. sp.
HOLOTYPE: USNM 1068555 About USNM , a dissected, slidemounted specimen.
TYPE LOCALITY: ALASKA: Adak Island: Little Thumb Creek , VII.1995 .
PARATYPES: U.S. National Museum, Washington DC. USNM 10685561068558 About USNM : Adak Island: Finger Creek , 27.VI.1995, 2 sectioned worms (1 transverse, 1 sagittal), 3 dissected .
OTHER MATERIAL: Adak Island : Finger Creek, 27. VI.1995, 2 sectioned worms (1 transverse, 1 sagittal); 5 dissected, partiallymature specimens and several immature whole mounts.
ETYMOLOGY: refers to the type locality in the Aleutian Islands , Alaska .
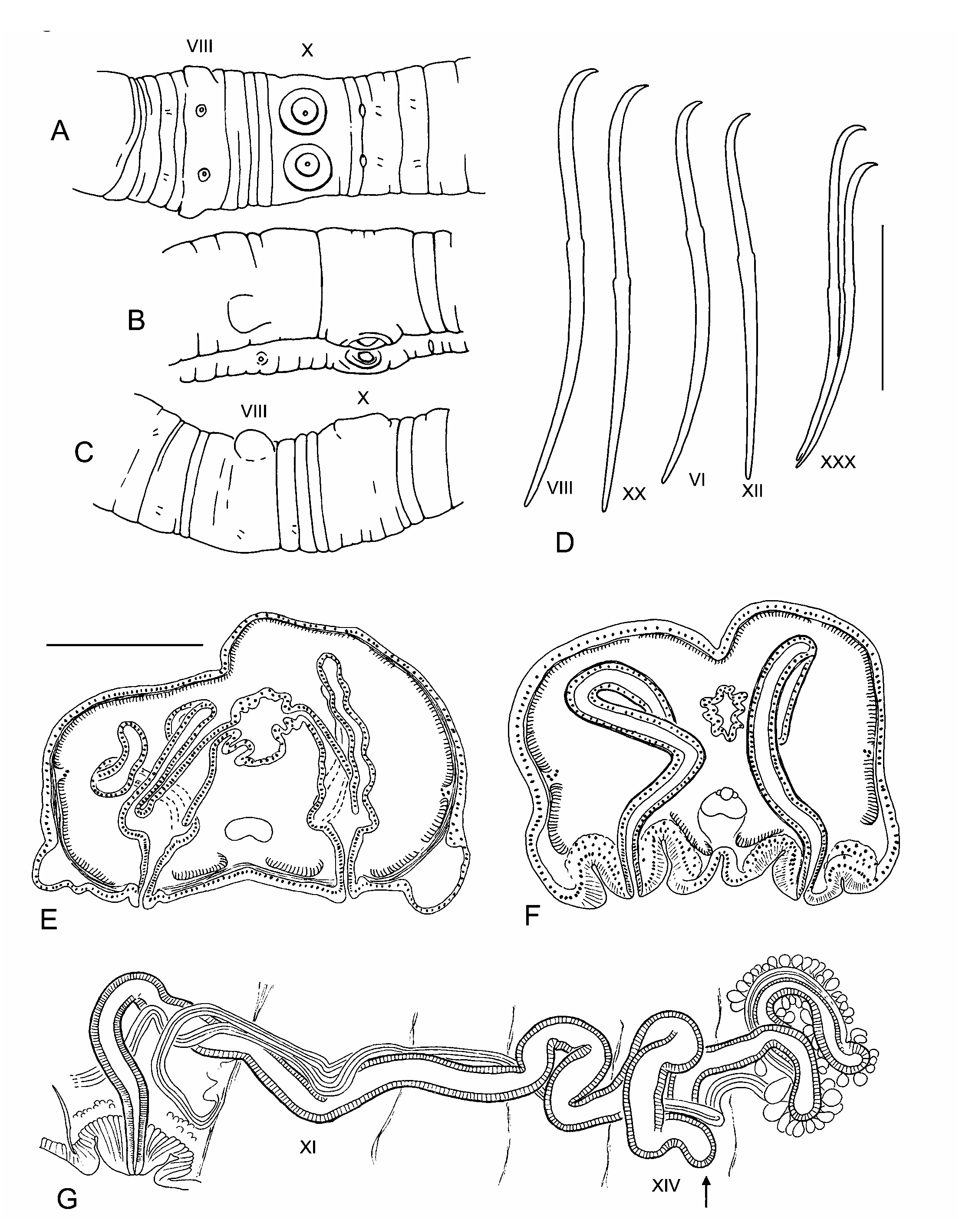
DESCRIPTION: Length of preserved specimens 18–25 mm, width 0.9–1.0 mm in X; 75–92 segments. Prostomium with a distinct proboscis, about 0.5 mm long by 0.05 mm diameter. Segmentation moderately developed; each segment posterior to II usually has a weak secondary annulus at about the anterior 1/4. Clitellum from IX or X to XIII. Chaetae simplepointed, sigmoid, with a moderate to strong distal hook; nodulus usually distal to midpoint, 35–45% of chaeta length from tip ( Fig. 1D View FIGURE 1 ), but sometimes median in posterior segments. Chaeta length 226–274 m in middle and posterior segments, dorsal chaetae may be slightly shorter than ventrals. Paired spermathecal openings on ventral chaetal lines, about midway between ventral chaetae and posterior septum in VIII. Ventrolateral margins of VIII may be expanded into blunt lobes, possibly claspers ( Figs. 1A–C, 1E View FIGURE 1 ). Male pores on ventral chaetal line, posterior to the chaetae in X, within short, raised rings of thickened epidermis ( Figs. 1A–C, 1F View FIGURE 1 , 2A View FIGURE 2 ). Female pores indistinct at 11/12, on ventral chaetal line.
Epidermis 14–17 m thick anteriorly, 20–50 m thick in clitellum. Longitudinal muscle layer of body wall 40–50 m thick in preclitellar segments; circular muscle layer 6–8 m. Longitudinal muscles divided at chaetal and lateral lines into bands which do not curl strongly inward at the edges ( Figs. 1E–F View FIGURE 1 ). Brain in the peristomium; major ventral nerves join in II. Pharynx in II–V, dorsal and ventral walls about equally thickened. Pharyngeal glands from IV to VI.
Ventral blood vessel divides in VII, branches connect to dorsal vessel in preclitellar segments via one pair of convoluted commissural vessels. No obvious lateral blood vessels posterior to X in the available material. Nephridia usually begin in XIII, paired, single, or absent in posterior segments. A small anteseptal funnel is followed by an irregular postseptal mass; a dorsal tubule loop approaches the dorsal blood vessel, a short tubule goes to an inconspicuous nephropore anterior to the ventral chaetal bundle, and another tubule may extend posteriorly into other segments.
Testes paired in X, ovaries paired in XI. Female funnels prominent at 11/12, pores intersegmental. Sperm sacs to as far as XVII in mature worms, egg sacs to as far as XXII. Vestigial sperm sacs usually paired, extending posteriorly from 8/9 and 9/10. Spermathecae paired in VIII, each pore may be surrounded by a broad, indistinctly raised area of slightly thickened epidermis; internally, pores may be surrounded by weakly developed accessory glands (not visible on most specimens). Spermathecae fusiform, 330– 600 m long, with about the middle third widened into an indistinct ampulla; entally, the ampulla narrows before it communicates with the gut ( Fig. 1E View FIGURE 1 ). One pair of unbranched diverticula joins ampulla near ectal end, on medial and lateral sides. Spermathecal diverticula tubular, 200–500 m long by 40–50 m in diameter; ental ends may be slightly expanded to about 60 m. Ectal duct and ampulla of spermatheca with outer muscle layer 5–10 m thick; muscle layer of diverticula 3– 4 m. Ectal duct 100–150 m long, 50–60 m wide at ampulla, but narrowed ectally, lined with columnar cells. Spermathecal sperm loosely arranged, but usually concentrated in ental ends of diverticula, with heads lined up along epithelium. Spermathecal ampulla, duct, and short diverticula develop before the ampulla joins the gut in partially mature specimens.
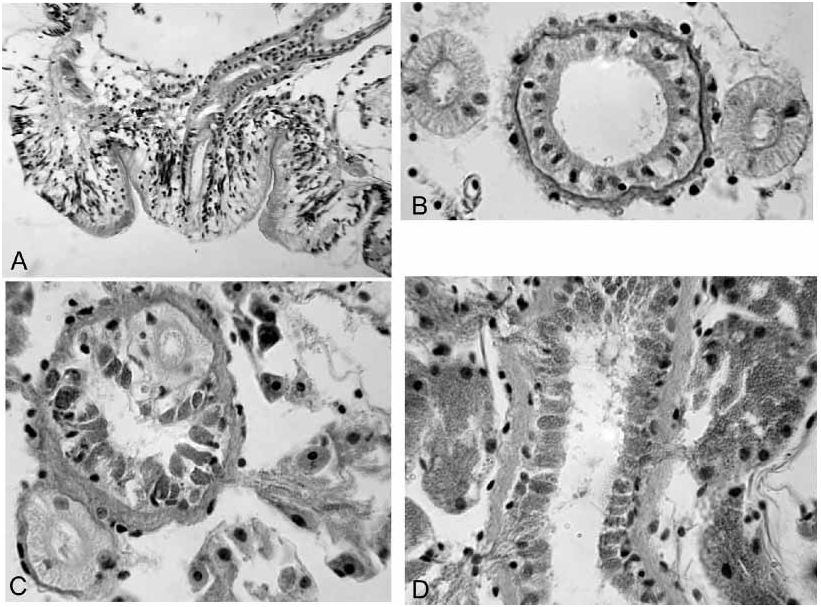
Male pores on low, roundedconical “penes” within shallow concavities surrounded by raised rings; the entire structure is an infolded pad of thickened epidermal cells ( Fig. 1F View FIGURE 1 ). Retracted penis to 130–220 m wide by 60–140 m tall; the entire pad is 250–430 m in diameter (but probably varies with contraction). Epidermal cells of pad and penis are elongate, with elongate nuclei; fibrous cell extensions form a dense ectal layer ( Fig. 2A View FIGURE 2 ). Unlike the surrounding clitellum, epidermal cells of pad do not appear to be glandular. Entally, the concave pad is surrounded by a thin layer of loose, somewhat granular cells, which sometimes appear as small, petiolate clumps, but may be interspersed with the epidermal cells in the penis ( Fig. 2A View FIGURE 2 ). Atria tubular, 2200 to 5000 m long, usually contorted, extending as far as XV ( Fig. 1G View FIGURE 1 ). Atrial diameter 50–100 m in ectal part, with muscle coat 2–5 m, and normal cuboidal epithelium ( Fig. 2B View FIGURE 2 ). Ental 1/4 to 1/2 of atrium 50–110 m in diameter, muscle layer 4–10 m thick, covered with petiolate, multicellular prostate glands ( Figs. 2C–D View FIGURE 2 ). Prostate glands 50 to 80 m long, highly granular, without distinct cell boundaries; narrow ends penetrate atrial muscle, forming clusters of extensions containing granular vacuoles among the atrial epithelial cells. Anterior male funnels cupshaped, to 180 m high, without sperm. Vasa deferentia ciliated, diameter usually 30–50 m ( Fig. 2B View FIGURE 2 ); anterior vas nonfunctional, but similar in size to or slightly narrower than the posterior. Both vasa deferentia join the atrial muscle coat near the middle of the prostatecovered portion or near the ental end; vasa travel under the muscle coat a short distance to enter the lumen slightly before the ental end ( Fig. 2C View FIGURE 2 ). Posterior male funnel with sperm; highly convoluted, 200 to 340 m in diameter, on 9/10. Posterior vas deferens enters X directly, without forming a loop into XI.
REMARKS: The two spermathecal diverticula, the modified male pores, the nonfunctional anterior vasa deferentia, and the tubular atria, having prostate bundles restricted to the ental part, appear to associate the new species with the “ Sutroa ” group of Nearctic species (equivalent to “Group 1” in Fend and Brinkhurst 2000; note that the group is no longer defined by a single median spermatheca). However, differences in morphological details suggests that R. aleutensis is distantly related to the more southern species. The structures surrounding the male pores could be interpreted as intermediate between the thick pads of R. brooksi Holmquist, 1976 and the distinctly invaginated, eversible penial bulbs of the Sutroa group (cf. Fend & Brinkhurst 2000: Figs. 77 vs. 22– 23). The structures may be eversible, although none was seen in an everted state, and distinct retractor muscles were not evident. Histologically, they appear as folded, thickened epidermis, rather than the mix of muscular and glandular cells seen in Sutroa group penial bulbs. Unfortunately, the poor histological condition of the available specimens limits the comparison.
Within the Sutroa group, R. aleutensis most closely resembles Rhynchelmis gilensis Fend & Brinkhurst, 2000 . Like R. gilensis , and in contrast to most Sutroa group species, the atrial muscle layer is not thickened in the ectal part, and both vasa deferentia enter the atria near the ental end. The paired spermathecae resemble the single, median spermatheca of R. gilensis , but lack the ectal “bulb” (cf. Fend & Brinkhurst 2000: Figs. 6, 46) and anterior pore position common to R. gilensis and all other species in the Sutroa group. The two spermathecal diverticula join the base of the ampulla, rather than the junction between the ectal duct and the ampulla.
The vasa deferentia are thicker than in any of the Nearctic Rhynchelmis s. lat. species, but do not appear to be glandular, or remarkably thickened (nearly as thick as the atria), as in the Palearctic Rhynchelmis s. str. (cf. Hrab 1982). Atria with prostate glands restricted to a relatively short portion of the atrium are also characteristic of Rhynchelmis brooksi and Rhynchelmis malevici (Sokolskaya 1983) , but those species have only a single vas deferens per atrium, and the middle portion of the atrium is histologically modified.
Rhynchelmis species typically have branched lateral blood vessels posterior to about VIII–X ( Cook 1971). The apparent lack of lateral blood vessels in R. aleutensis View in CoL may be an artifact of poor fixation, but bloodvascular systems appear to be reduced within closely related Rhynchelmis species groups (e.g. Rhynchelmis alaskana Holmquist, 1976 View in CoL vs. Rhynchelmis saxosa Fend & Brinkhurst, 2000 View in CoL ). The nephridia resemble other Rhynchelmis View in CoL in having a postseptal duct loop that approaches the dorsal blood vessel ( Fend & Brinkhurst 2000).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Rhynchelmis aleutensis
| Fend, Steven V. 2005 |
R. aleutensis
| Fend 2005 |
Rhynchelmis saxosa
| Fend & Brinkhurst 2000 |
Rhynchelmis alaskana
| Holmquist 1976 |
Rhynchelmis
| Hoffmeister 1843 |