
Zeanillus Jeannel 1937
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4196.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:026D88BD-86C0-4811-814A-CB5CACB3104F |
|
DOI |
https://doi.org/10.5281/zenodo.6089664 |
|
persistent identifier |
https://treatment.plazi.org/id/8D378784-891F-FFCE-21CA-3E70FC0FFC5E |
|
treatment provided by |
Plazi |
|
scientific name |
Zeanillus Jeannel 1937 |
| status |
|
Zeanillus Jeannel 1937 View in CoL
Zeanillus Jeannel, 1937: 277 View in CoL (type species Anillus phyllobius Broun, 1893 View in CoL , by original designation)
Recognition. The members of this genus are distinguished from the other New Zealand representatives of Anillina by the following combination of characters: eyes absent; head with long fronto-lateral carinae; antennae of moderate length; prosternal process tapering to the blunt apex; pronotum cordiform with rounded posterior angles; elytra lacking oblique longitudinal grooves, with lateral margins slightly denticulate and with slightly dehiscent apices with rounded sutural angles; pygidium exposed at least in apical half; 1st and 3d discal setae of elytra always present; umbilicate series of 9 pores: the longest setae in the 2nd, 6th and 9th pore positions, 7th, 8th and 9th pores not aligned, virtually forming obtuse isosceles triangle with 8th pore shifted towards disc, 8th and 9th pores of umbilicate series much closer to each other than the 7th pore is to the 8th. Among characters listed above only the shape of prosternal process might be treated as an autapomorphy of the genus, as it is not seen in any other New Zealand Anillina except Hygranillus View in CoL , which is known to me by description only. Apart from the shape of the prosternal process, members of Zeanillus View in CoL can be distinguished from representatives of other New Zealand genera of Anillina as follows. Developed fronto-lateral carinae, antennae of moderate length, discal setae in basal half of elytra, and exposed pygidium separate Zeanillus View in CoL members from those of the endogean Hygranillus Moore. Rounded View in CoL posterior angles of pronotum and smooth elytra distinguish representatives of Zeanillus View in CoL from those of species Pelodiaetus Jeannel View in CoL and Pelodiaetodes Moore. The View in CoL umbilicate series of nine pores, dehiscent apices of elytra, and exposed pygidium separate members of Zeanillus View in CoL from those of Nesamblyops View in CoL .
Description. Size. SBL range 1.41–2.01 mm.
Habitus. Body form from slightly convex to subdepressed, ovoid or subparallel ( Fig. 11 View FIGURE 11 ).
Color. Body rufotestaceous or testaceous, appendages testaceous.
Microsculpture. Dorsal microsculpture of polygonal sculpticells with isodiametric mesh pattern over the dorsal surface. Development of microsculpture varies on different body parts. On head and disc of pronotum, microlines less pronounced, fine, while on elytra microlines very distinct, forming well-defined sculpticells.
Luster. Body surface shiny.
Macrosculpture. Body surface sparsely and finely punctate.
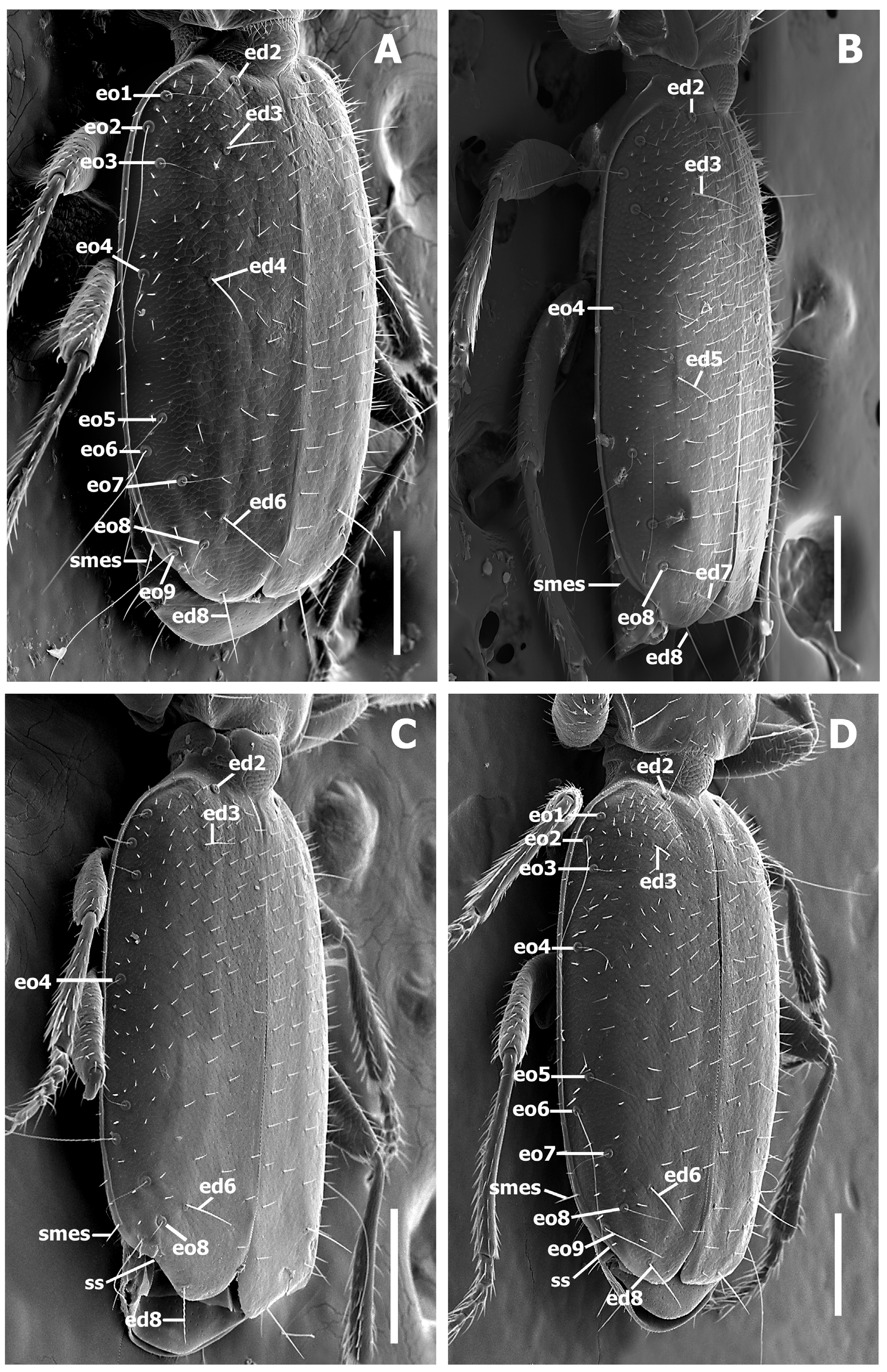
Vestiture. Body surface covered with sparse yellowish short setae. Vestiture of elytra short (around one-third length of discal setae). Last marginal elytral setae anterior to subapical sinuation vary in length among different group of species (cf. Fig. 8 View FIGURE 8 A versus D, smes).
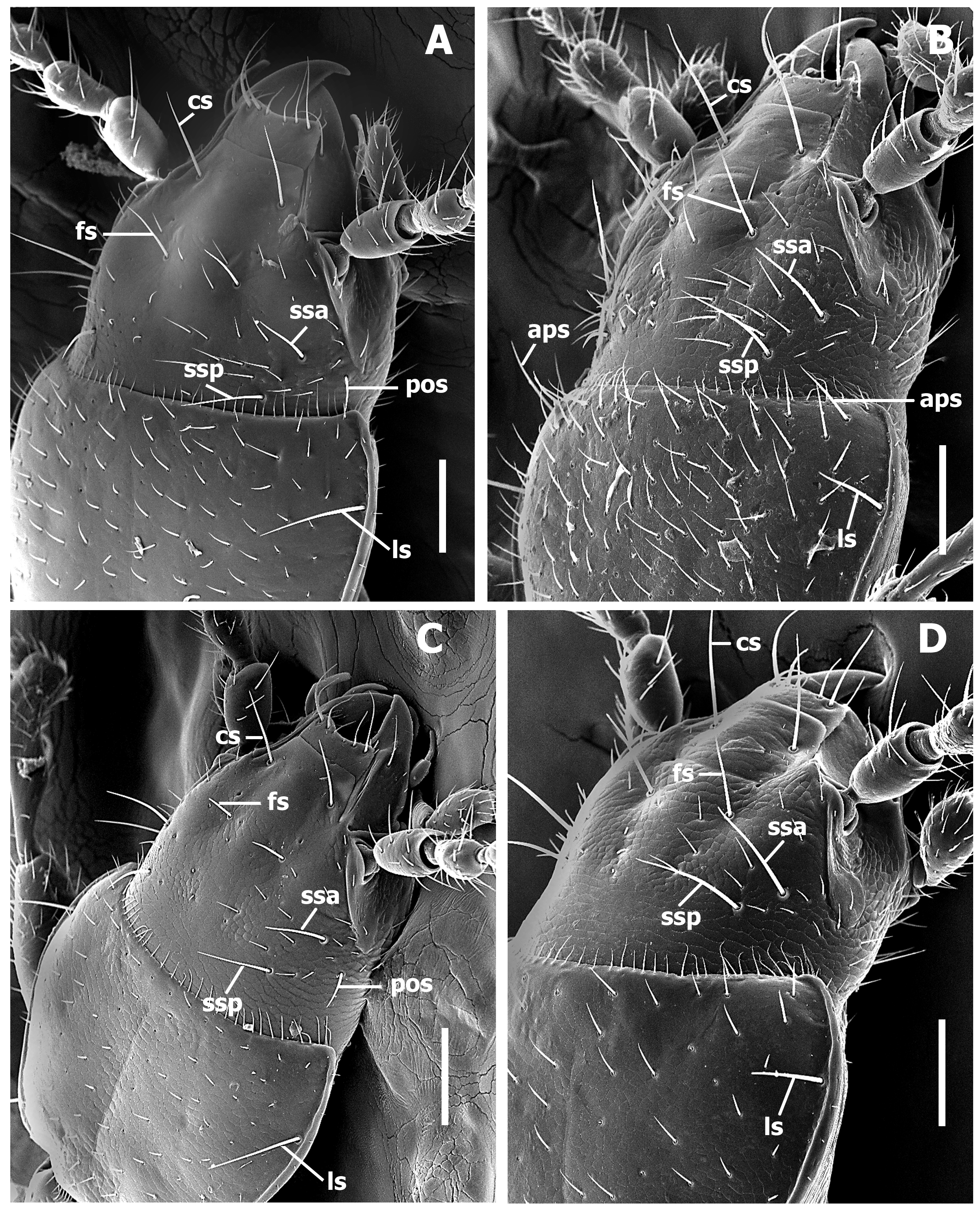
Fixed setae. Primary head setae include a pair of clypeal (cs), a pair of frontal (fs), two pairs of supraorbital (ssa and ssp), and additionally one pair of postorbital (pos) setae ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ) in two groups of species. Mentum with two pairs of long primary (paramedial and lateral) setae ( Fig. 4 View FIGURE 4 , pms, lms). Submentum with three pairs of long primary setae (lss, prss) and few additional shorter setulae ( Fig. 4 View FIGURE 4 ). Maxilla with long stipetal and palpiferal setae. Pronotum with two long primary lateral setae (midlateral, ls, and basilateral, bs) on each side ( Fig. 5 View FIGURE 5 ) and in one species with distinct apicolateral (aps) setae ( Fig. 2 View FIGURE 2 B). Elytra, depending on group of species, either with 3 ( Fig. 8 View FIGURE 8 A–B) or 2 discal setae ( Fig. 8 View FIGURE 8 C–D), with scutellar (ed2) and apical (ed8) setae. Last three (7th, 8th, and 9th) pores (eo7, eo8, and eo9) of umbilicate series not aligned, with 8th pore shifted towards the disc and virtually forming an obtuse isosceles triangle, The 8 th and 9 th pores of umbilicate series closer to each other than the 7 th pore is to the 8th, the longest setae in the 2nd, 6th, and 9th pore position ( Fig.8 View FIGURE 8 A). Fifth visible sternite of male with two and of female with four setae along the posterior margin.
Head ( Fig. 1 View FIGURE 1 ). Anterior margin of clypeus (cl) straight. Frontal area flat without tubercle medially near frontoclypeal suture. Fronto-lateral carinae distinct and long (fcc).
Eyes. Eyes absent.
Antennae. Submoniliform, with 11antennomeres, extended to about posterior margin of pronotum. Antennomeres 1 and 2 elongate, of equal length and 1.4–1.5 times longer than antennomere 3, which is only slightly elongate and 1.1–1.2 times longer than antennomere 4. Antennomeres 4 to 10 globose, last antennomere conical and 1.6–1.7 times longer than penultimate antennomere.
Labrum ( Fig. 1 View FIGURE 1 ). Labrum (lb) transverse with shallowly concave, entire anterior margin with six setae apically, increasing in size from the central pair outwards.
Mandibles ( Fig. 3 View FIGURE 3 ). Similar to Pelodiaetodes ( Sokolov 2015) . Right mandible with distinct anterior (art) and posterior retinacular (prt), small terebral (tt), distinct premolar (pm), and molar (mt) teeth. Left mandible with distinct terebral (tt), premolar (pm) and molar (mt) teeth only.
Maxillae. Maxillary palps with basal trianguloid cardo, and stipes with dorsal and ventral lobes, dimerous galea, and standard lacinia. Palpus ( Fig. 4 View FIGURE 4 ) with short 4th palpomere (mp4), 0.15–0.2 length of palpomere 3 (mp3).
Labium ( Fig. 4 View FIGURE 4 ). Labium with short acute or blunt mental tooth (mt) and in one case without it ( Fig. 4 View FIGURE 4 D); mentum (m) and submentum (sm) split, with mental-submental suture (ms) and in one species labium fused, without mental-submental suture ( Fig. 4 View FIGURE 4 D). Glossal sclerite (gsc) with shortly lobed paraglossae (pg), with two setae medio-apically.
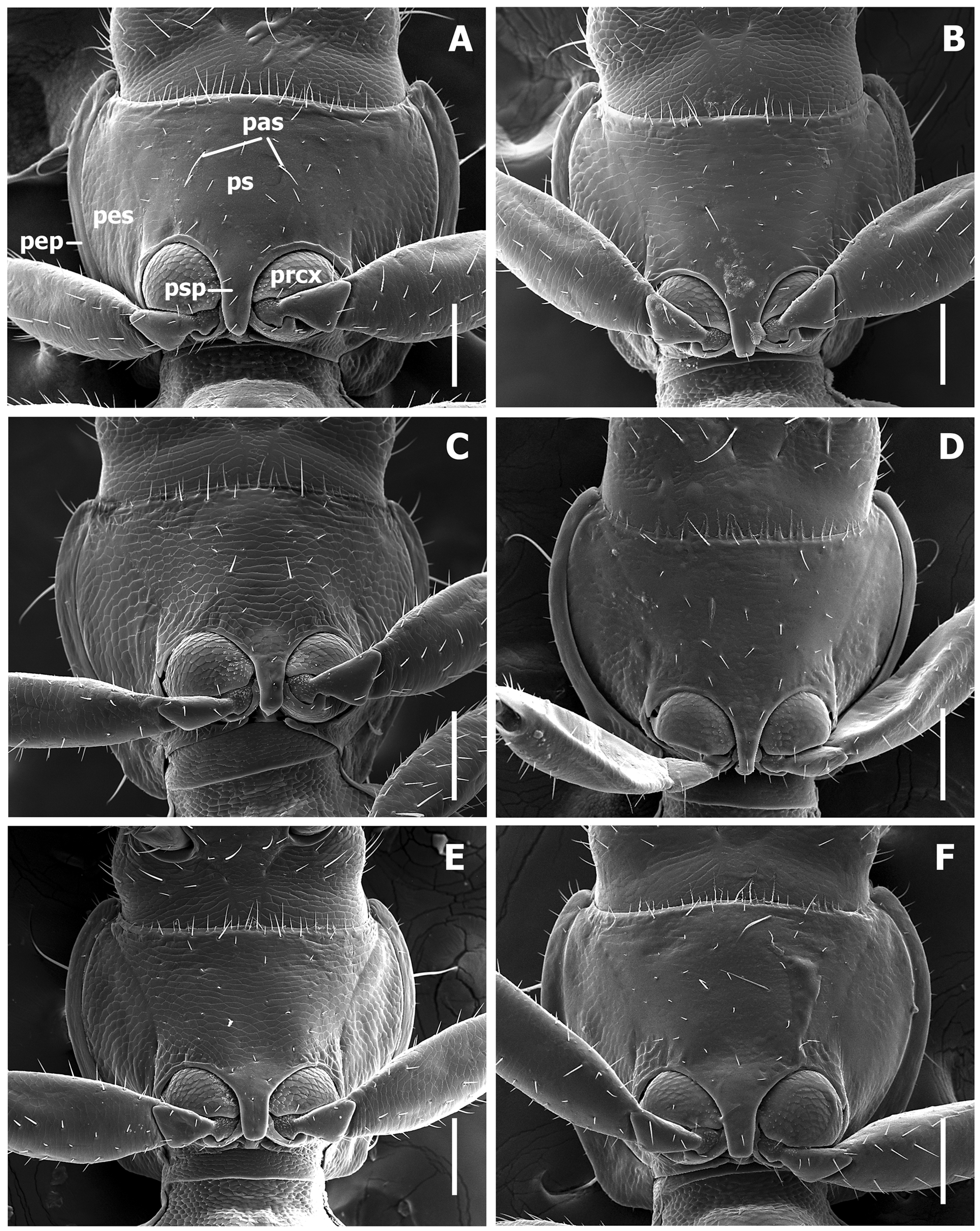
Prothorax ( Fig. 5 View FIGURE 5 ). Pronotum cordiform, moderately convex, either rectilinear constricted posteriorly or slightly sinuate anterior to posterior angles, with narrow marginal gutter (mg). Posterior margin of pronotum shallowly concave medially. Anterior angles almost rounded, barely projected forward. Posterior angles rounded, nearly effaced, basilateral seta inserted far anterior to angles. Widths across anterior margin much greater than between posterior angles at the level of basilateral setae (WPa/WPp varies from 1.15 to1.51 among species). Prosternum ( Fig. 6 View FIGURE 6 ) slightly projected at the anterior margin medially, there with a group of longer setae relative to other prosternal vestiture, also with a pair of long ambulatory sensor setae (pas) at the middle of sclerite. Prosternal intercoxal process (psp) unmargined, slightly tapering apically and obtusely truncate at apex.
Scutellum ( Fig. 5 View FIGURE 5 B). Externally visible, triangular, with obtuse apex.
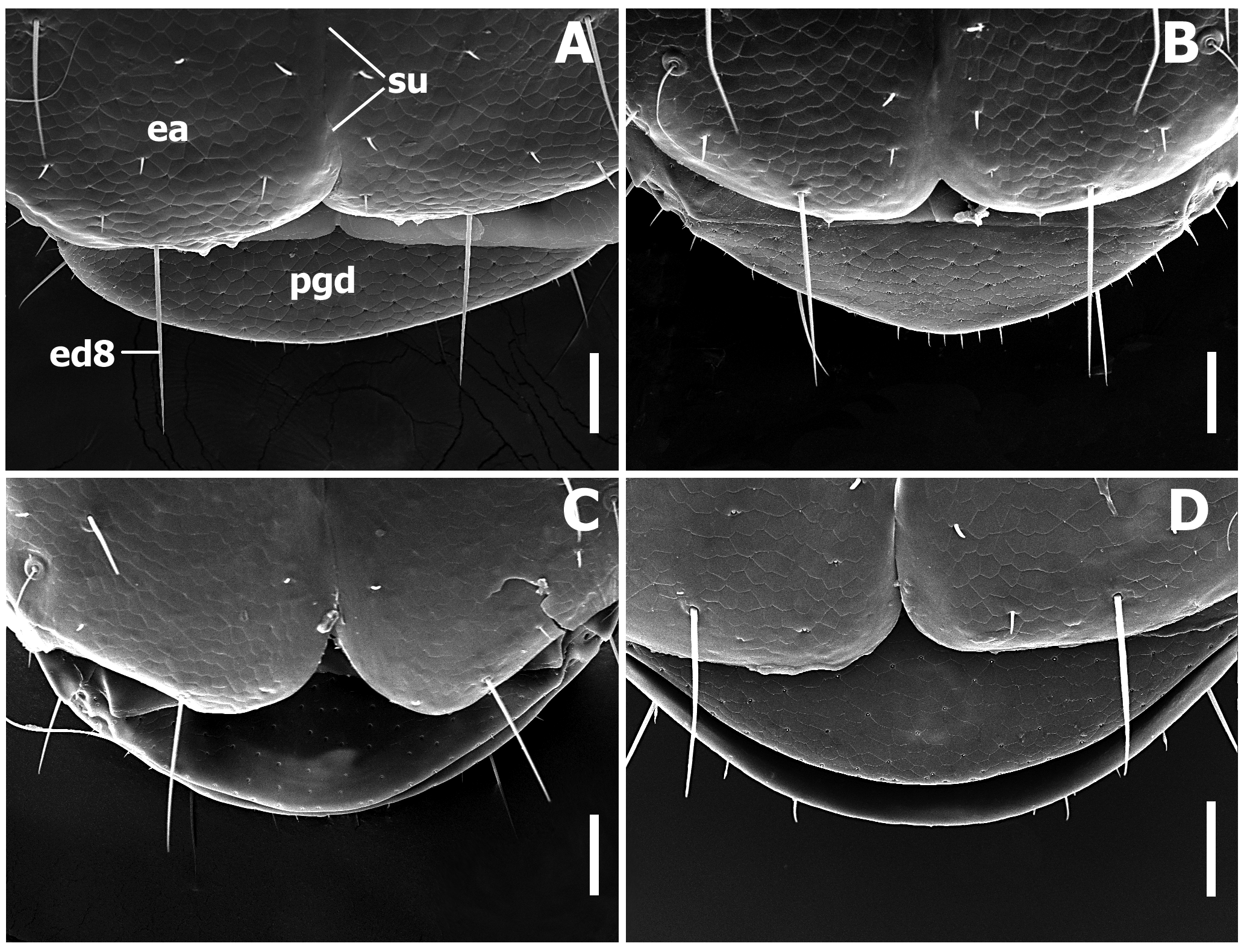
Elytra ( Fig. 8–9 View FIGURE 8 View FIGURE 9 ). Elytra subdepressed, relatively short (LE/SBL from 0.47 to 0.55 among species) without visible interneurs and without longitudinal grooves. Humeri rounded, to form oblique angle with longitudinal axis of body. Elytral basal margination lacking ( Fig. 5 View FIGURE 5 A–C). Apical half of elytra with shallow subapical sinuation ( Fig. 8 View FIGURE 8 , ss). Sutural angle of elytron rounded, making apices of elytra slightly dehiscent ( Fig. 9 View FIGURE 9 ).
Hind wings. Absent.
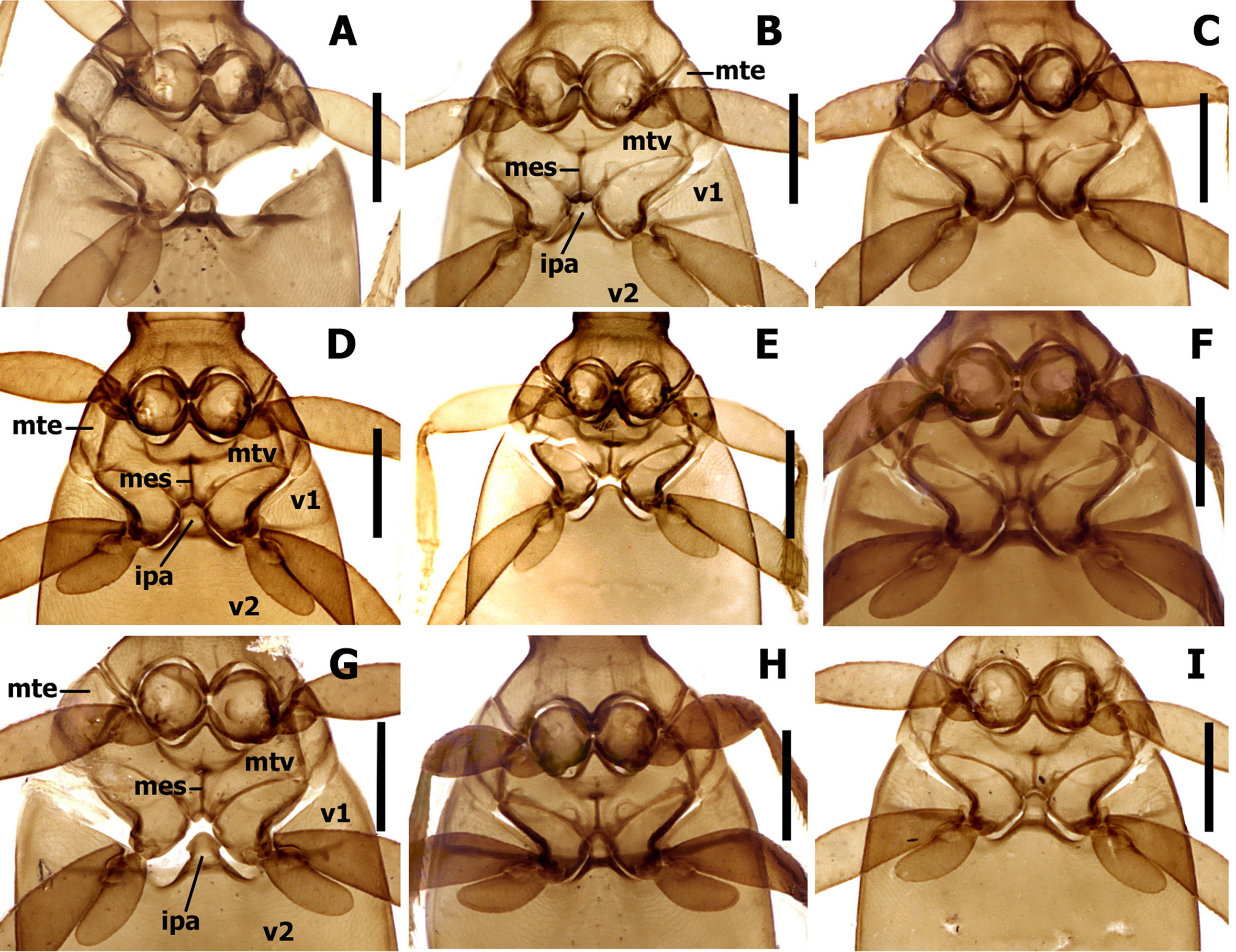
Pterothorax ( Fig. 7 View FIGURE 7 ). Metaventrite (mtv) short, distance between meso- and metacoxae about 0.5–0.6 diameter of mesocoxa. Metanepisternum (mte) short, subquadrate, with anterior and outer margins of approximately equal length. Metendosternite (mes) cross-shaped with lateral arms. Lateral arms either V-shaped with divergent branches or U-shaped with almost subparallel branches ( Fig. 7 View FIGURE 7 D–E).
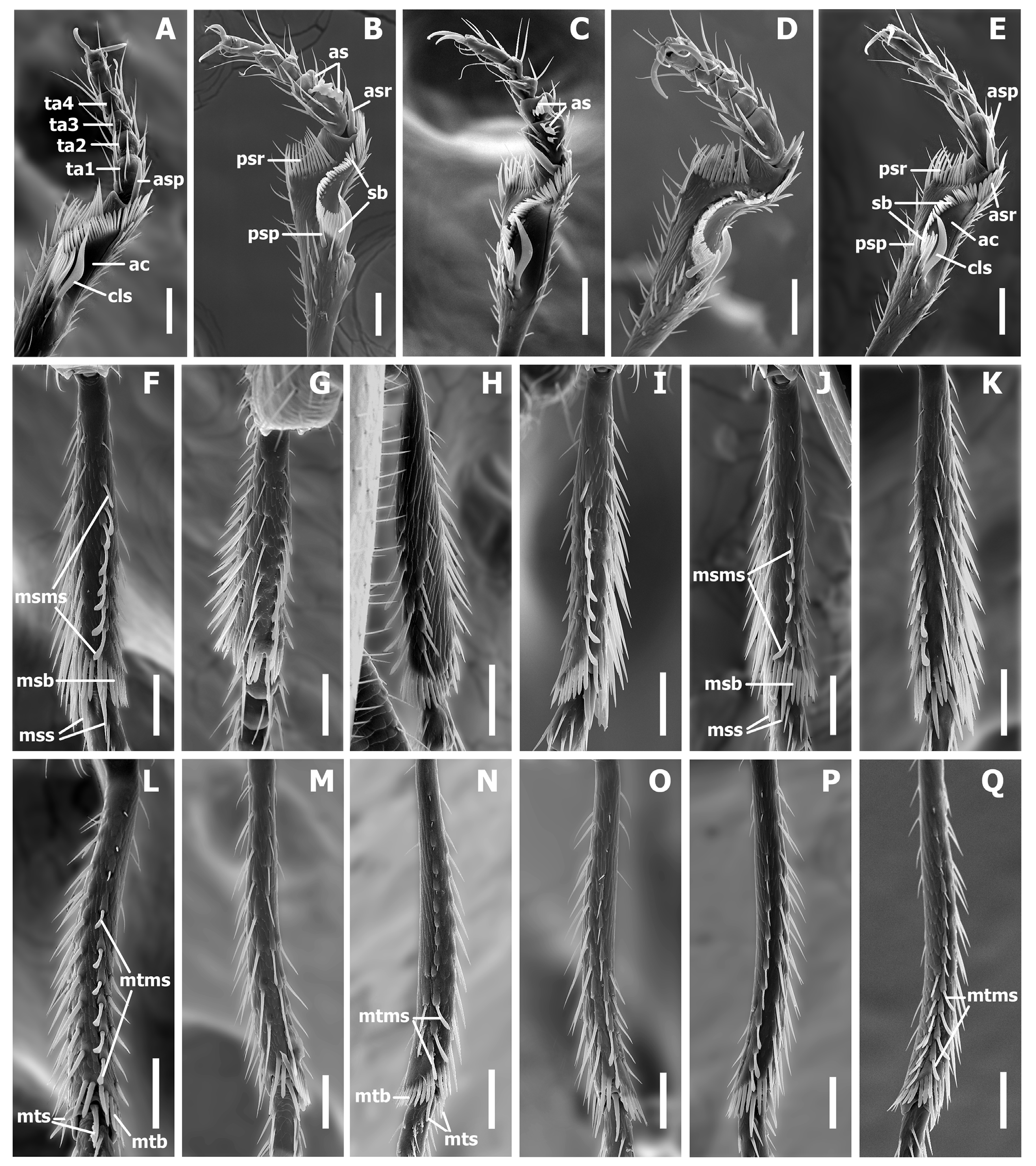
Legs ( Fig. 10 View FIGURE 10 ). Legs of moderate length, not elongate. Prothoracic legs of males with first 2 tarsomeres (ta1–2) moderately dilated apico-laterally with two rows of oval articulo-setae (as) ( Stork 1980) on the ventral surface ( Fig. 10 View FIGURE 10 B–C). Protibiae with antenna cleaner of type B ( Hlavac 1971), with both anterior (asr) and posterior (psr) apical setal rows and concave apico-lateral notch ( Fig. 10 View FIGURE 10 A–E). Length of anterior spur (asp) slightly smaller than length of 1st tarsomere (ta1). Profemora moderately swollen. Mesotibiae ( Fig. 10 View FIGURE 10 F–K) with one long row of modified ventral setae (msms) at apical half, with two terminal spurs (mss) and tibial brush (msb). Metafemora unmodified, metatibiae ( Fig. 10 View FIGURE 10 L–Q) with one row of modified ventral setae (mtms) in apical half, with two terminal spurs (mts) and tibial brush (mtb). Number of metatibial modified setae varied among species. Tarsi pentamerous, last and 1st tarsomeres are the longest, 2–4 tarsomeres of equal length on the tarsi of all legs, 1st tarsomere shorter than combined length of 2–4 tarsomeres. Tarsal claws simple, untoothed.
Abdominal ventrites. Five visible abdominal ventrites: 2nd ventrite longest, 2.8–3.5 times longer than 3rd or 4th, 3rd and 4th equal in length; the last, 5th, 1.4–1.6 times longer than 4th. Intercoxal process of 2nd ventrite of moderate width, triangularly constricted anteriorly ( Fig. 7 View FIGURE 7 , ipa).
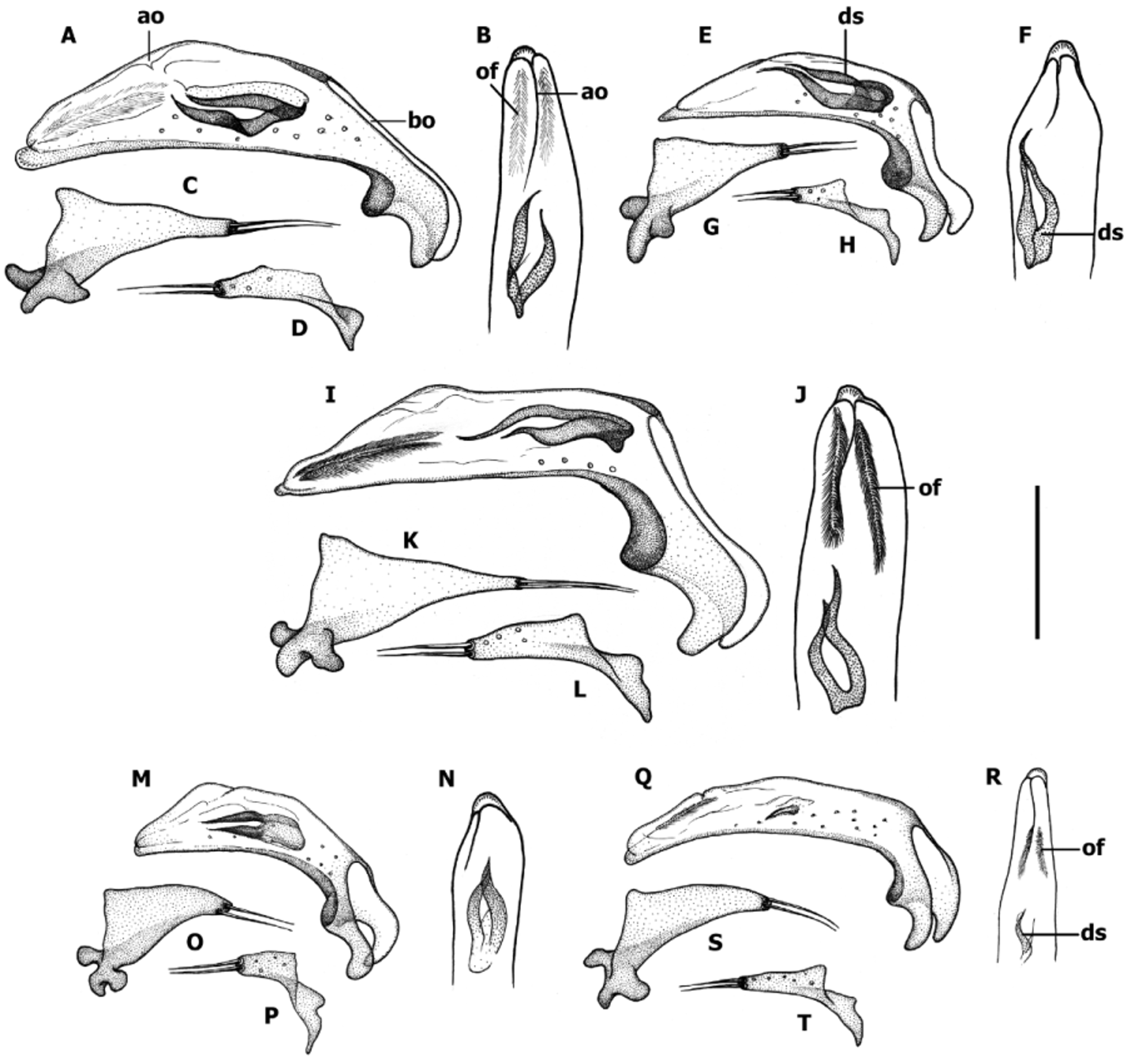
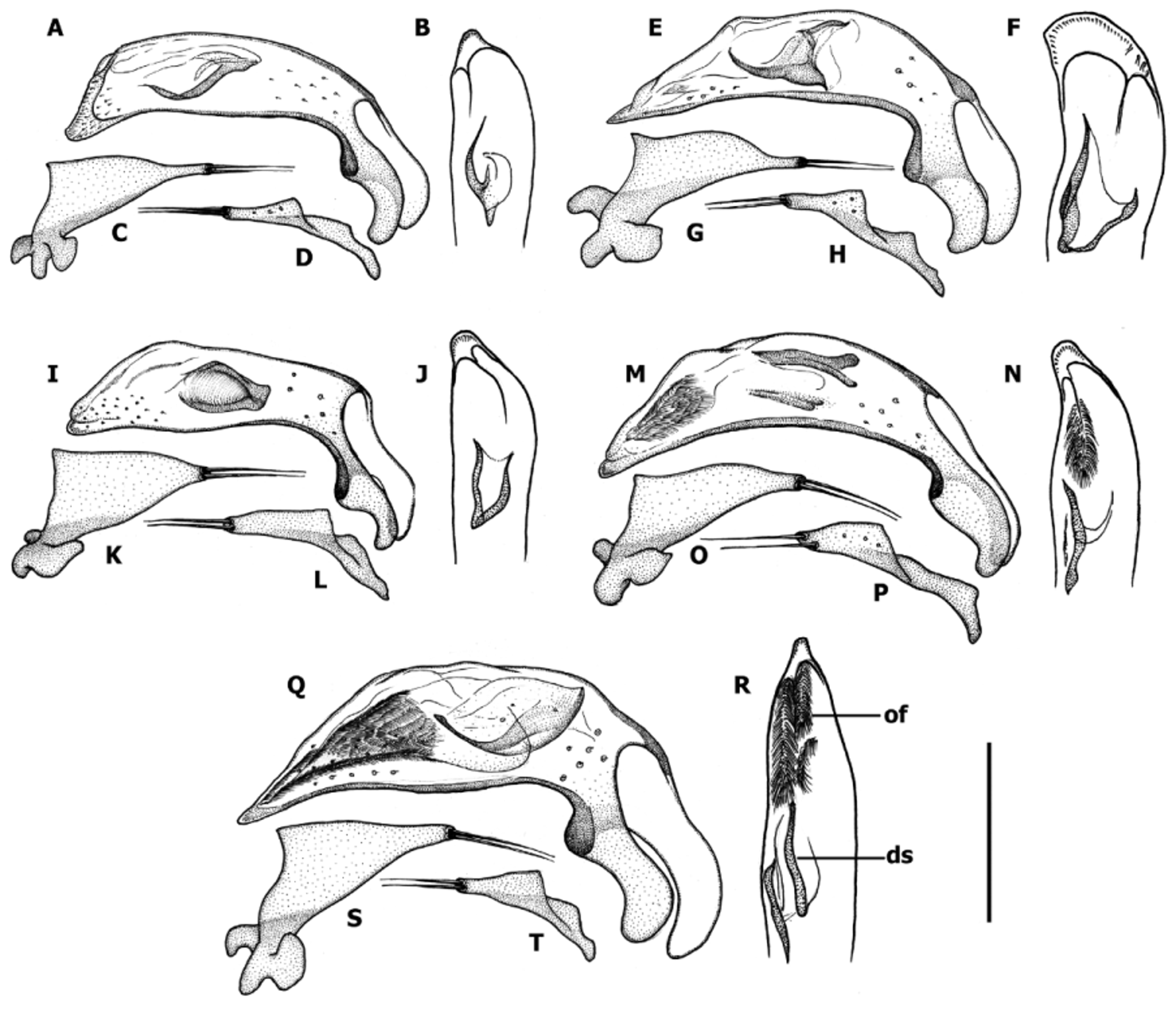
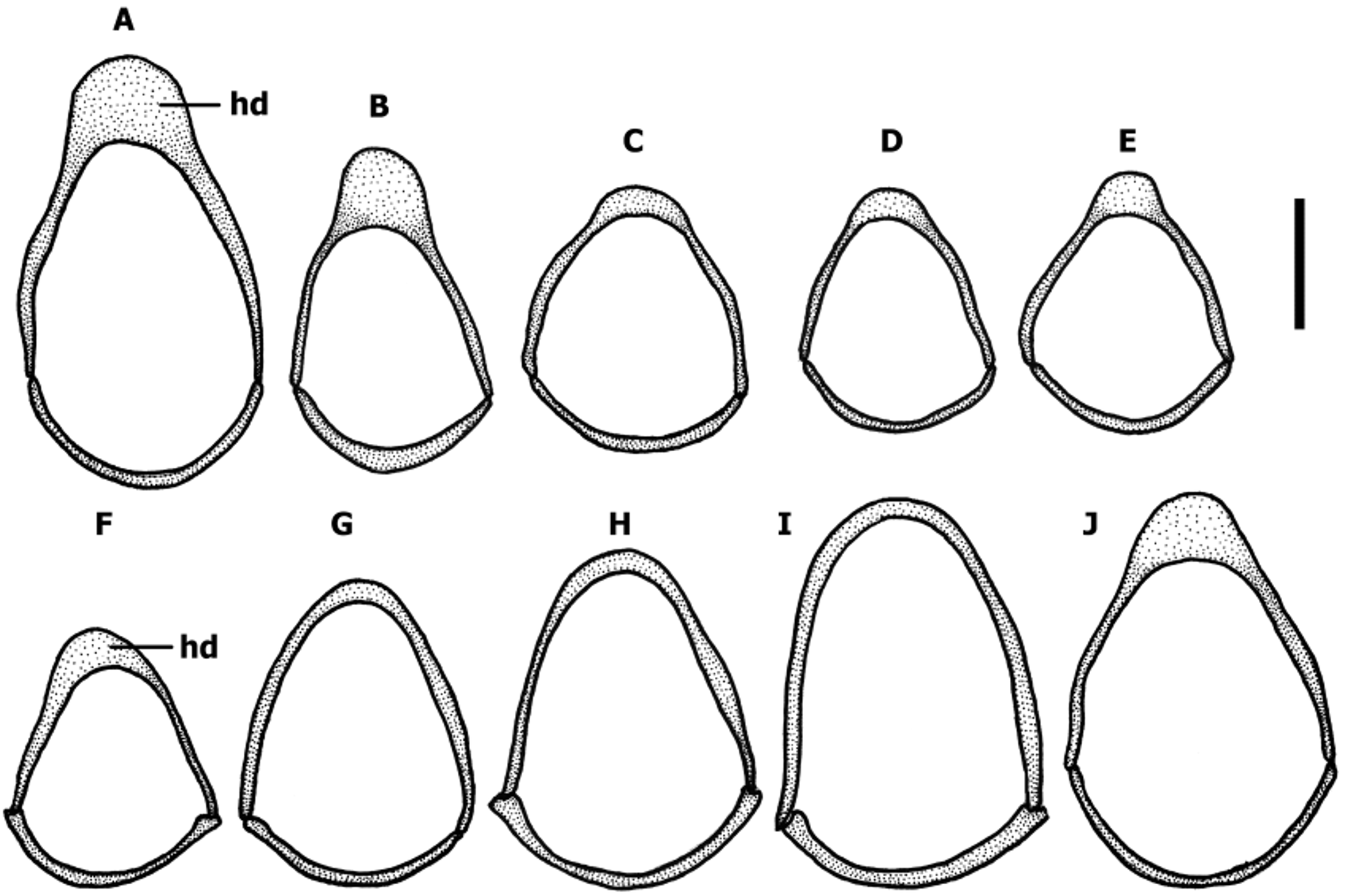
Male genitalia ( Fig. 12–14 View FIGURE 12 View FIGURE 13 View FIGURE 14 ). Median lobe of aedeagus anopic, elongate, slightly twisted, and moderately arcuate ( Fig. 12–13 View FIGURE 12 View FIGURE 13 ). Apex of median lobe typically unmodified, except two species with enlarged apex. Internal sac with two copulatory sclerites of various size, represented either by two separate structures or by two sclerites united basally, in the last case sclerites acutely attenuated apically. Some species have ostial fields at apical orifice. Additional spines of internal sac are absent. Parameres bisetose. Left paramere large and broad either evenly tapered to apex or attenuated and subparallel proximal to setal attachment, right paramere short. Ring sclerite ( Fig. 14 View FIGURE 14 ) ovoid either with transverse broadly rounded handle-like extension (hd) varying in length among species or simple with no traces of enlargement.
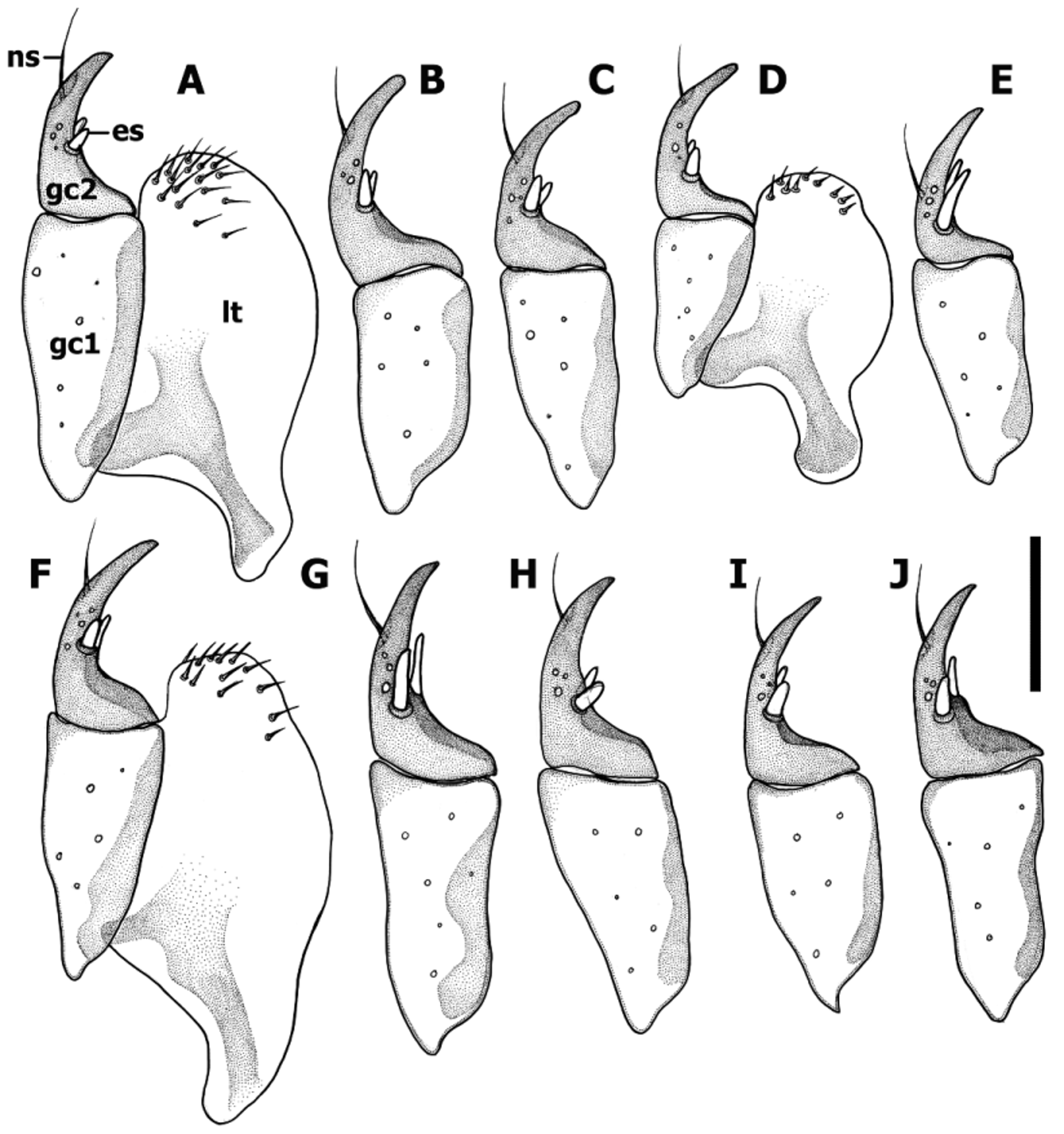
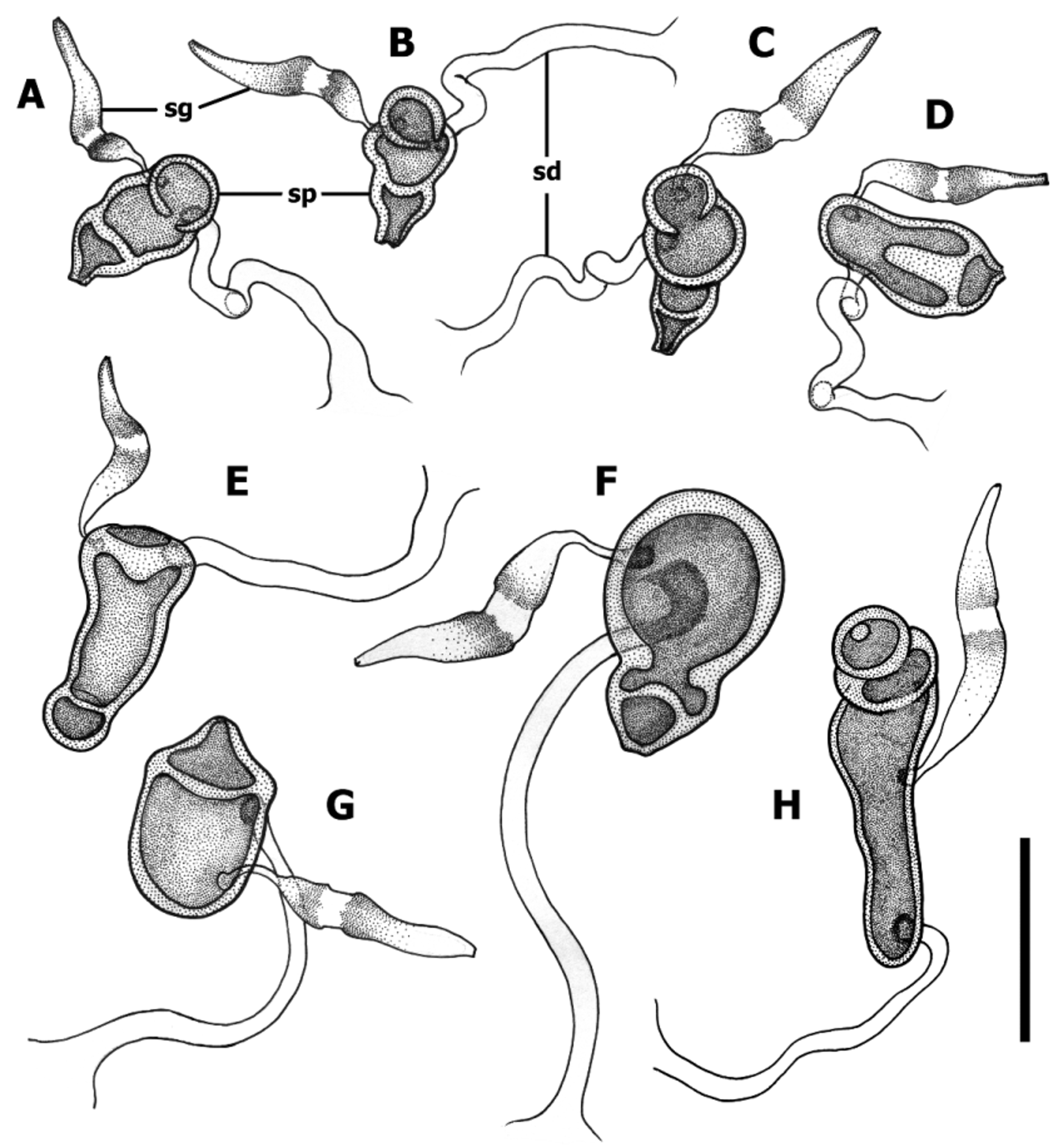
Female internal genitalia ( Fig. 15–17 View FIGURE 15 View FIGURE 16 View FIGURE 17 ). Gonocoxite 1 asetose ( Fig. 15 View FIGURE 15 , gc1). Gonocoxite 2 falciform (gc2), 1.6–1.8 times longer than its basal width, moderately curved, with ensiform (es) and apical nematiform (ns) setae. Length of ensiform setae varies among species. Laterotergite (lt) with 8–16 setae. Spermatheca (sp) sclerotized ( Fig. 16–17 View FIGURE 16 View FIGURE 17 ), of different size (cf. Fig. 17 View FIGURE 17 A versus C), length and shape, with 2–4 inner compartments, making traditional naming of spermathecal parts impossible. Spermathecal duct typically short, except Z. carltoni with very long and coiled duct ( Fig. 17 View FIGURE 17 A).
Included taxa. The genus comprises four subgenera and ten species: nominotypical subgenus includes Z. phyllobius (Broun) , Z. punctigerus (Broun) , and Z. nunni , sp.n.; subgenus Brounanillus, subgen.n., includes Z. pallidus (Broun) ; subgen Nunnanillus, subgen.n., includes Z. pellucidus , sp.n.; and subgenus Otagonillus, subgen.n., includes Z. brouni , sp.n., Z. carltoni , sp.n., Z. lescheni , sp.n., Z. montivagus , sp.n., and Z. nanus , sp.n.
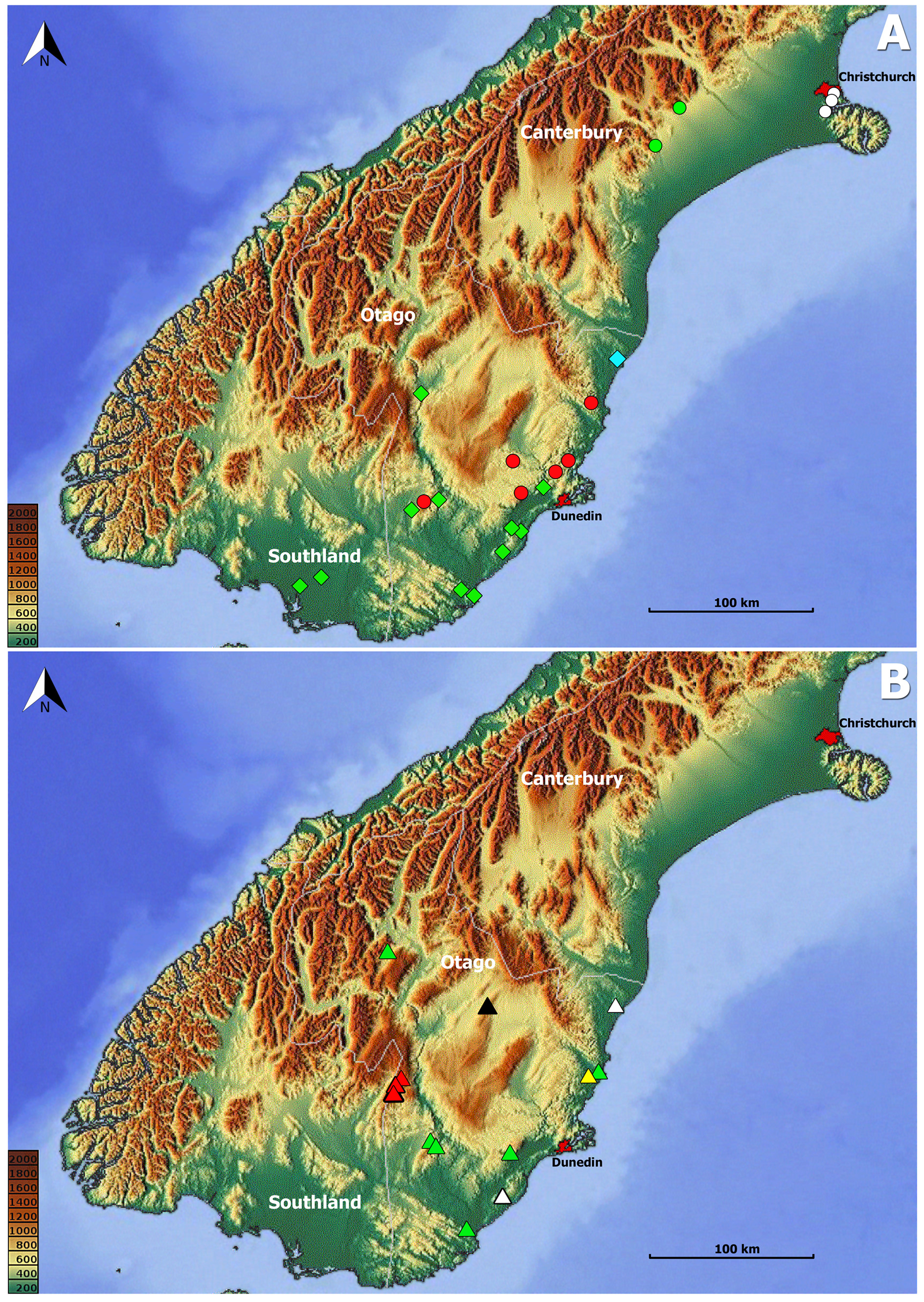
Geographical distribution. The species of Zeanillus are known from three regions of the South Island of New Zealand ( Fig. 18 View FIGURE 18 ): Canterbury, Otago, and Southland . The majority of the species inhabit Otago (8 species), two species are confined to Canterbury and three (two shared with Otago) are known from Southland .
Way of life. According to the label information, specimens of Zeanillus were collected mostly from washed soil samples, sometimes in a combination with turf or moss. Only a few specimens were collected from litter and one specimen was collected under a stone after rain. Collections were made in a vast spectrum of habitats: from subalpine tussocklands to lowland and coastal forests, including Podocarp-kamahi, Podocarp-broadleaved, and beech-broadleaved communities. Beetles were collected during all months throughout the year, except March.
Relationships. The varied state of mental tooth (absent or present) together with the structure of the umbilicate series: nine pores with the longest seta in 2nd, 6th, and 9th pore position, formally allow different members of Zeanillus to be placed within two different subdivisions of Anillina: either the Zeanillus - or the Microtyphlus phyletic lineages sensu Jeannel (1963) and Giachino (2008). The Zeanillus phyletic lineage embraces all New Zealand genera and is restricted in distribution to New Zealand only. Among the New Zealand genera the putative closest relatives of the members of Zeanillus are the representatives of Nesamblyops . Both genera share distinct paraglossae, rounded posterior angles of pronotum, and smooth elytra, distinguishing them from any other species of the New Zealand anillines. However, difference in the structure of the prosternal process and in the structure of the male and female reproductive organs suggest only a remote relationship between the two genera. Representatives of the Microtyphlus phyletic lineage inhabit South Europe, Africa with Madagascar and Australia. Some of them possess the same combination of external characters as the members of Zeanillus , like noncrenulated basolateral margins of pronotum, rounded posterior pronotal angles, dehiscent elytra, and exposed pygidium, thus superficially resembling the latter. Among these taxa are some species of Microtyphlus Linder ( Jeannel 1963) from Europe, the members of Caeconannus Jeannel (Jeannel 1963, Giachino 2015) from South Africa, the single representative of Bulirshia Giachino (Giachino 2008) from Magadascar and one species of Pseudillaphanus Giachino (Giachino 2005) from Australia. However, the absence of paraglossae in all of the mentioned taxa immediately distinguish their members from Zeanillus . So, it is likely that external similarities among the taxa listed above reflect convergences rather than relationships and true relatives of Zeanillus among Anillina outside New Zealand can be determined only after thorough molecular phylogenetic analysis. Until then, it is best to consider the members of Zeanillus of autochthonous origin with obscure relationships outside New Zealand.
Classification. All species were arranged in four clusters based on peculiarities of chaetotaxy of the head, pronotum, and elytra, primarily on the state of postorbital and discal elytral fixed setae, allowing placement of all species into unambiguous groups. Additionally, states of the labium and mental tooth are involved in distinguishing one group. Traditionally these last characters have been used for the supraspecific ranking of Anillina, mainly for distinguishing new genera (for Microtyphlus -phyletic lineage cf. Giachino 2008; Jeannel 1963). I prefer to propose these groups of species as subgenera, mainly to conserve the generic status of this otherwise morphologically welldefined and apparently monophyletic group.
The key provided below allows identification of adults of the Zeanillus






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Trechinae |
|
Tribe |
Bembidiini |
Zeanillus Jeannel 1937
| Sokolov, Igor M. 2016 |
Zeanillus
| Jeannel 1937: 277 |