
Marbefia carthyi ( Hamond, 1968 ), 2009
|
publication ID |
https://doi.org/10.3897/zookeys.23.168 |
|
publication LSID |
lsid:zoobank.org:pub:A447D3B6-5387-44B6-AC1B-A05988000E43 |
|
DOI |
https://doi.org/10.5281/zenodo.3790666 |
|
persistent identifier |
https://treatment.plazi.org/id/8F78FF1D-160C-CB4F-C380-1FFBFC0ED081 |
|
treatment provided by |
Plazi |
|
scientific name |
Marbefia carthyi ( Hamond, 1968 ) |
| status |
comb. nov. |
Marbefia carthyi ( Hamond, 1968) , comb. n.
Figs 1–6
Pseudonychocamptus carthyi Hamond, 1968
Type locality. England, Norfolk, Hunstanton; in washings of colonies of Hartlaubella (as Laomedea ) gelatinosa ( Pallas, 1766) ( Hydrozoa, Campanulariidae ) attached to concrete lumps found in little shallow pools under the pier; for associated fauna see Hamond (1968: 176).
Material examined. ( 1) Holotype. ♀ dissected on one slide (reg. no. NHM 1967.10.2.2), from type locality, collected on 2 April 1957 by R. Hamond;
(2) one ♀ dissected on 12 slides, one ♁ dissected on 9 slides, one ♀ and one ♁ used for SEM examination, and two ♁♁ in 70% ethanol; all specimens collected from Station MB.17 (53°10.34΄ N, 00°56.34΄ E) on 6 May 1992 by R .
Figure |. Marbefia carthyi ( Hamond, 1968) , comb. n.: A habitus ♀, dorsal B habitus ♀, lateral C habitus ♁, dorsal.
Hamond (reg. nos. NHM 1993.312–321); depth 12–13 m; fine sand with high silt content;
(3) one ♀ (reg. no. NHM 1998.588) re-identified by F. Fiers in 1998, found among Pseudonychocamptus koreni ( Boeck, 1873) specimens (reg. no. NHM 1967.10.31 –70) collected during the 1966 University of London Sub-Aqua Club expedition to the Isles of Scilly; Great Britain Rock, off St Mary’s; algal debris at LWN, among bryozoans ( Cellaria , Lepralia ) and hydroids ( Sertularia ), 27 m; July 1966 ( cf. Wells 1970: 267).
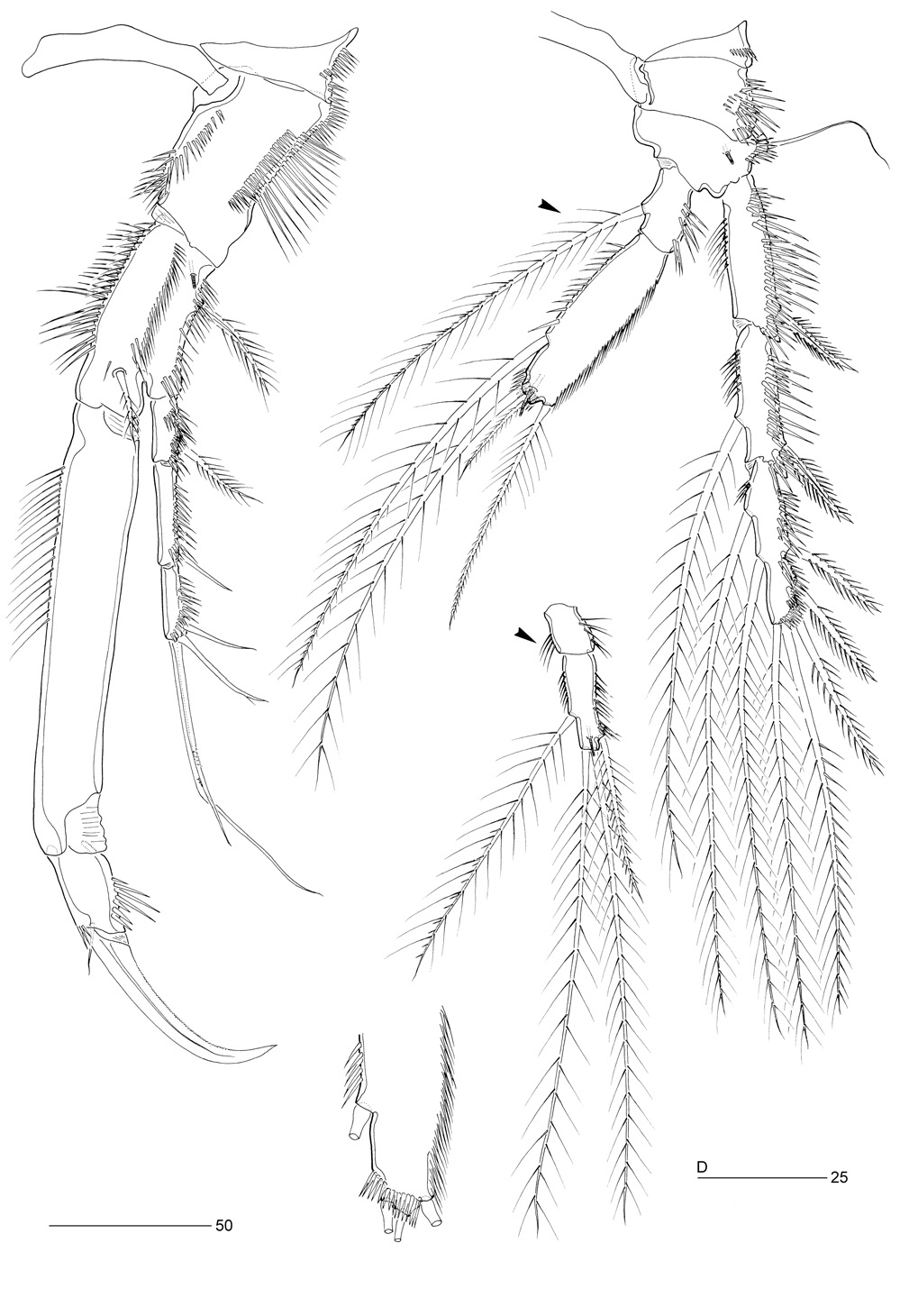
Description. Female. Body (Figure 1A, B). Total body length 696–738 µm (n=2; mean 717 µm). Largest width at posterior margin of cephalic shield ( 174 µm). Urosome gradually tapering posteriorly (Figure 1A). Rostrum triangular ( Figure 2C View Figure 2 ), with pair of sensilla near apex; midventral tube-pore in subapical position; completely defined at base. Cephalothorax with posterior margin fringed with small spinules; pleural areas well developed and rounded with lobate posterolateral angles; entire surface covered with tiny spinules as illustrated in Figure 1A, B; sensilla and few pores present as illustrated in Figure 1A, B. Pedigerous somites covered with minute spinules. All prosomites without defined hyaline frills; posterior margins fringed with small spinules. Body constricted between individual somites. Urosome (Figures 1A, B; 2A) 5-segmented, comprising P5-bearing somite, genital double-somite and 3 free abdominal somites. Abdominal half of genital double-somite and second abdominal somite with lateral lobate extensions. All urosomites with surface ornamentation consisting of small spinules dorsally and laterally; ventral surface largely naked except for few rows of spinules and paired tube-pores; ventral hind margin with large spinules laterally and small spinules medially ( Figure 2A View Figure 2 ). Hyaline frills of urosomites not distinct. Genital double-somite (Figures 1A; 2A) with transverse, surface ridge dorsally and laterally, indicating original segmentation; completely fused ventrally. Genital field ( Figure 2G View Figure 2 ) with medium-sized copulatory pore (arrowed in Figure 2G View Figure 2 ) located in median depression; gonopores fused medially forming single genital slit covered on either side by operculum derived from sixth leg; P6 with small protuberance bearing 2 bare setae; small triangular process located on medial side adjacent to protuberance. Anal somite ( Figure 2B View Figure 2 ) with spinular ornamentation and pair of dorsal tube-pores; anal operculum well developed, bearing row of minute spinules and flanked by pair of sensilla; anal opening bordered by well developed frill bearing long setular extensions.
Caudal rami ( Figure 2A, B View Figure 2 ). Short, cylindrical, as long as wide; each ramus with 7 setae: seta I subventral, bare and shortest; setae II and III bare; setae IV and V bipinnate, fused basally, and with predesigned fracture planes [seta V presumably longest but incomplete in all specimens examined including holotype, cf. Hamond (1968: Fig. 3 View Figure 3 )] ; seta VI bipinnate; seta VII tri-articulate at base. Each ramus with spinules on outer dorsal surface; additional spinular ornamentation present along inner and outer margins and around ventral and dorsal hind margins; long tube-pore present near ventral hind margin.
Antennule ( Figure 2D View Figure 2 ). Seven-segmented, slender; with well developed sclerite around base of segment 1; several rows of long spinules present along posterior margins of all segments except for apical segment, and along anterior margins of segments 1–4. Segments 1–2 without spinous processes. Segment 2 largest; segment 4 with aesthetasc fused basally to seta and arising from distinct pedestal. Armature formula: 1-[1], 2-[7 + 1 pinnate], 3-[7], 4-[1 + (1 + ae)], 5-[1], 6-[2], 7-[7 + acrothek]. Apical acrothek consisting of small aesthetasc fused basally to 2 bare setae.
Antenna ( Figure 3A View Figure 3 ). Three-segmented, comprising coxa, allobasis and free 1-segmented endopod. Coxa small, with 2 rows of spinules. Allobasis not elongate; without distinct surface suture marking original segmentation but with membranous insert around base of exopod; with one abexopodal bipinnate seta in distal half. Exopod small, twice longer than width, with 4 well developed pinnate setae (2 laterally, 2 apically), distal lateral and inner distal ones with finer spinules; one row of coarse spinules on anterior surface. Endopod shorter than allobasis; lateral armature arising in distal half, consisting of one bare seta flanked by 2 strong pinnate spines; apical armature consisting of 2 strong spines and 3 geniculate setae (one geniculate seta fused basally to short seta). Endopod with 2 rows of long spinules laterally and 2 transverse hyaline frills subapically.
Labrum. Well developed; spinular ornamentation on anterior surface as in Figure 2E View Figure 2 .
Mandible ( Figure 3B View Figure 3 ). With well developed gnathobase bearing several multicuspidate teeth around distal margin and one pinnate spine at dorsal corner as in original description ( Hamond 1968: Fig. 6 View Figure 6 ). Palp ( Figure 3B View Figure 3 ) small; endopod and exopod fused to basis, represented by small peduncles bearing 3 plumose and one pinnate seta(e), respectively. Basal armature represented by one plumose seta.
Paragnaths ( Figure 2F View Figure 2 ). Strongly developed paired lobes with medially directed hair-like setules, separated by medial lobe covered by dense pattern of short setules.
Maxillule ( Figure 3C View Figure 3 ). Praecoxa with few long spinules around outer margin; arthrite strongly developed, with a tube-seta on anterior surface and 8 spines/setae around distal margin; one transverse row of long spinules on posterior surface, and one row of small spinules along inner margin of arthrite. Coxa with cylindrical endite bearing a naked seta and a curved, pinnate spine; with spinular row on anterior surface. Basis with cylindrical endite bearing 2 naked setae and a pinnate spine; with several spinular rows on anterior surface. Endopod completely incorporated into basis, forming cluster of 3 plumose setae; exopod 1-segmented, with 2 plumose setae and several rows of fine spinules on surface.
Maxilla ( Figure 3D View Figure 3 ). Syncoxa with 3 endites; with a row of long spinules along outer margin, several rows of spinules on posterior surface, and several spinules along inner margin; praecoxal endite small and cylindrical, with one bare seta; both coxal endites with a strong pinnate spine and 2 pinnate setae with subapical tubular extension; proximal coxal endite with one row of spinules anteriorly. Allobasis drawn out into strong, slightly curved, distally pinnate claw; accessory armature consisting of 2 pinnate and 2 naked setae. Endopod represented by one bipinnate and 3 naked setae, surrounded by spinules at base.
Maxilliped ( Figure 3E View Figure 3 ). Syncoxa elongate, with 2 plumose setae and two rows of spinules. Basis with one row of spinules along outer margin, palmar margin completely bare. Endopod drawn out into long claw; with one short accessory seta anteriorly, and one tube-pore posteriorly.
Swimming legs. P1–P4 ( Figures 4A, B View Figure 4 ; 5 View Figure 5 A–C). With wide intercoxal sclerites and well developed praecoxae. Praecoxae, coxae and bases with anterior rows of surface spinules as figured. Exopods 3-segmented, endopods 2-segmented.
P1 ( Figure 4A View Figure 4 ). Coxa large; with several spinular rows and patches as figured. Basis with bipinnate seta on distal pedestal near insertion of endopod, long setules along inner margin and one bipinnate spine and several spinules along outer margin; anterior tube-pore near articulation with coxa. Exp-1 with one bipinnate seta; exp-2 with one naked outer seta; exp-3 with 2 virtually naked and 2 geniculate setae. Enp-1 1.8 times as long as exopod, with long spinules along proximal half of inner margin; enp-2 with one strong, minutely denticulate claw, and one small naked seta; several spinules along outer margin and around inner distal corner.
P2–P4 ( Figures 4B View Figure 4 ; 5A, B View Figure 5 ). Coxae and bases with spinular rows along outer margin and on anterior surface; basis with tube-pore on anterior surface; outer margin of basis with bipinnate spine (P2) or naked seta (P3–P4); exopodal and endopodal segments with elaborate spinular/setular ornamentation as figured. P4 outer exopodal spines with long spinules. P2–P4 endopods with short enp-1 and long, inflated enp-2; outer margin of enp-2 weakly chitinized, with 2 parallel rows of flimsy setular extensions, possibly surrounding glandular structure ( Figure 5B View Figure 5 ). P3–P4 enp-2 with tube-pore near distal margin. P2 enp-2 twice times longer than enp-1; endopod reaching to distal margin of exp-2; exp-3 longest. P3 enp-2 2.5 times longer than enp-1; endopod reaching to distal margin of exp-2; exp-3 longest. P4 enp-2 2.6 times longer than enp-1, posterior distal margin with spinular row ( Figure 4D View Figure 4 ); endopod reaching to about distal margin of exp-2; exp-3 subequal to exp-1. Outer distal seta of P4 enp-2 clearly shorter than others; inner seta of enp-1 very long (arrowed in Figure 4B View Figure 4 ) and much longer than in P3 enp-1 (arrowed in Figure 5C View Figure 5 ). Spine and setal formulae of swimming legs as for genus.
Fifth pair of legs ( Figure 6E View Figure 6 ). Baseoendopods not medially fused, with distinct medial extension. Exopod and baseoendopod discrete, each with pattern of spinules as figured; spinular pattern on exopod denser than on baseoendopod. Baseoendopod forming long, outer setophore bearing basal seta and rows of spinules; with 3 tubepores on anterior surface; endopodal lobe just extending to proximal area of exopod, with 2 apical and 3 lateral bipinnate setae, proximal and inner middle ones minutely pinnate. Exopod elongate, about 3 times longer than wide; with one naked terminal seta, one bipinnate inner seta, and one bare and 3 pinnate setae along outer margin; terminal seta arising from short cylindrical process.
Egg-sac. Number of eggs and arrangement unknown.
Description. Male. Body (Figure 1C). Body length 570–587 µm (n=3; mean 579 µm). Largest width at posterior margin of cephalic shield ( 144 µm). Urosome narrower than prosome. Prosome (Figure 1C) 4-segmented, comprising cephalothorax and 3 free pedigerous somites. Cephalothorax with smooth posterior margin; pleural areas well developed and rounded, with lobate posterolateral angles; whole surface covered with tiny spinules as in ♀. Rostrum as in ♀. Pedigerous somites covered with minute spinules as in ♀. Urosome (Figure 1C) 6-segmented, comprising P5-bearing somite, genital somite and 4 abdominal somites. All urosomites with surface ornamentation consisting of tiny spinules dorsally and laterally; ventral surface smooth except for few rows of small spinules as in ♀; ventral posterior margin with large spinules laterally and minute spinules medially. Hyaline frills of urosomites not distinct.
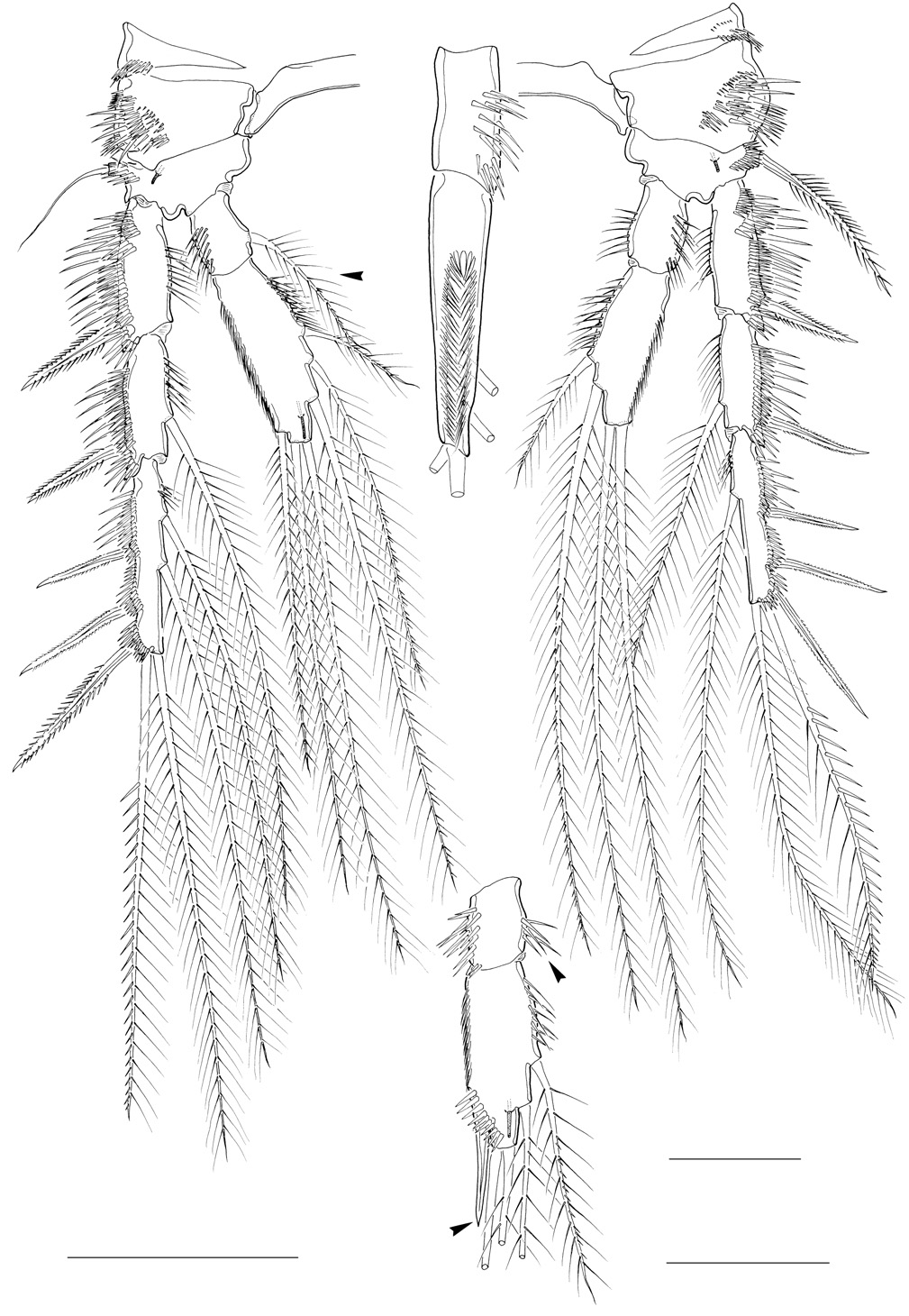
Antennule ( Figure 6 View Figure 6 A-C). Eight-segmented; subchirocer with geniculation between segments 5 and 6. Segments 1 and 2 with several rows of long spinules along anterior and posterior margins; without protuberances. Segment 3 ( Figure 6B View Figure 6 ) with few spinules along anterior margin. Segment 4 represented by small U-shaped sclerite around anterior margin (see insert in Figure 6B View Figure 6 ). Segment 5 swollen. Segment 8 triangular ( Figure 6C View Figure 6 ). Segmental homologies: 1-(I), 2-(II–VIII), 3-(IX–XII), 4-(XIII), 5-(XIV–XX), 6-(XXI–XXII), 7-(XXIII), 8-(XXIV–XXVIII). Armature formula: 1-[1], 2-[1 pinnate + 8], 3-[8], 4-[2], 5-[9 + 1 pinnate + 1 modified + (1 + ae)], 6-[3 modified spinous elements], 7-[1], 8-[8 + acrothek]. Apical acrothek consisting of minute aesthetasc and 2 naked setae.
Swimming legs. P2–P4 ( Figures 4C View Figure 4 ; 5D View Figure 5 ). Shape and surface ornamentation of intercoxal sclerites and protopods as in ♀. P2–P3 enp-2 with double row of setular extensions along outer lateral margin as in ♀ (absent in P4; Figure 4C View Figure 4 ). No sexual dimorphism on P2. Endopodal segments of P3–P4 modified.
P3 ( Figure 5D View Figure 5 ). Exopod as in ♀. Endopod 2-segmented, modified, not reaching to distal margin of exp-2. Enp-1 with several spinules on inner distal margin (arrowed in Figure 5D View Figure 5 ); inner seta absent. Enp-2 produced distally into short, straight, spiniform apophysis (homologous with outer spine of enp- 2 in ♀; arrowed in Figure 5D View Figure 5 ) surrounded at base by row of coarse spinules.
P4 ( Figure 4C View Figure 4 ). Exopod as in ♀. Endopod relatively shorter than in ♀. Enp-1 with several spinules along inner margin (arrowed in Figure 4C View Figure 4 ); inner seta absent; outer lateral margin without setular extensions found in ♀. Enp-2 twice longer than enp-1; both apical setae about equally long and longer than in ♀.
Fifth pair of legs ( Figure 6D View Figure 6 ). Baseoendopods fused medially, with spinular patch medially. Baseoendopod with large setophore bearing outer basal seta; endopodal lobe vestigial bearing one pinnate and one bare seta; with one tube-pore along medial margin, and one tube-pore near articulation with exopod; with few coarse spinules at base of setophore. Exopod about twice as long as maximum width; with one inner, one apical and 2 outer setae; all setae pinnate except for distal outer one being smooth; anterior surface with several large spinules.
Sixth pair of legs ( Figure 6D View Figure 6 ). Asymmetrical; represented on both sides by small plate (fused to ventral wall of supporting somite on one side; articulating at base and covering gonopore on other side; dextral and sinistral configurations observed); outer distal corner produced into cylindrical process bearing several spinules, one bipinnate inner and one naked outer seta.
Discussion. Hamond’s (1968) original description of the female of Pseudonychocamptus carthyi is detailed by contemporary standards. Our redescription revealed some minor observational errors: (1) the entire body is covered by fine spinules instead “… a few long limp hairs, widely scattered here and there”; (2) the third antennulary segment has 7 setae instead of 5; (3) the distal lateral and inner distal setae of the antennary exopod have fine spinules instead of membranous laminae; (4) the antennary endopod has an additional subdistal spine and the outermost geniculate seta is fused at the base to a minute seta; (5) the basis and coxal endite of the maxillule have an ad- ditional seta; (6) the maxilla has a unisetose praecoxal endite and the endopod 4 setae instead of 2; (7) the P1 displays distinct spinule rows along the outer margin of the praecoxa and coxa, and enp-2 has a minute accessory seta at the base of the claw; (8) the outer spines of P2–P3 exp-2 and -3 are bipinnate instead of smooth; and (9) the caudal ramus has 7 setae (seta I was overlooked).
The discovery of the male of P. carthyi provides conclusive evidence not only for the inclusion of this species in Lee and Huys’ (1999) PWPH -clade (grouping the genera Pilifera , Pseudonychocamptus , Weddellaophonte and Heteronychocamptus ) but also for its exclusion from the genus Pseudonychocamptus . The sexual dimorphism in swimming leg armature (the inner seta of P3–P4 enp-1 being absent in the male) unequivocally links this species to the PWPH -clade (Table 1). The sexually dimorphic P4 endopod, being 2-segmented in the female but reduced to a minute bisetose segment in the male, is the most significant apomorphy of the genus Pseudonychocamptus , however, no such reduction is displayed by the male of P. carthyi . This species also displays the most primitive armature on P2–P5 within the PWPH -clade (Table 1), lending additional support to a different generic assignment. The only other genus exhibiting two inner setae on P3–P4 exp-3 is Weddellaophonte , however, in P. carthyi two additional setae are found on P3 enp-2 (formula 321). Pseudonychocamptus carthyi is here fixed as the type of a new genus, Marbefia , gen. n., which can be defined by the following autapomorphies: (1) P2–P4 enp-2 (except P4 enp- 2 in ♁) inflated, with weakly chitinized outer margin bearing two parallel rows of flimsy setular extensions, possibly surrounding a glandular structure; a similar but non-homologous structure is found on the P4 endopod of species of Corbulaseta Huys & Lee, 2000 ( Huys and Lee 2000; Gómez and Boyko 2006); (2) P4 with sexually dimorphic setation on enp-2 (outer distal seta reduced in ♀, very long in ♁); and (3) antennule ♀ with spinules along posterior margins of segments 1–6.
Fiers (1992: 213) hinted at a relationship between Pseudonychocamptus and the monotypic genus Harrietella T. Scott, 1906 ( type: Laophonte simulans T. Scott, 1894a ) but provided no supporting evidence for this claim. Presumably his assertion was adopted from Lang (1948: 1449) who recognized a similar relationship based on the shared more or less dorsoventrally flattened body and the 2-segmented P3 endopod and 1-segmented P4 endopod in the male, in addition to the overall similarity in the maxilliped, P1 and male P3 exopod. Most Pseudonychocamptus species, however, do not have a dorsoventrally depressed body and the general habitus is considerably different from that of Harrietella simulans (T. Scott 1894a; Sars 1920b). In Pseudonychocamptus the P4 endopod is suppressed during development in the male but not in the female, resulting in different segmentation and setation patterns, whereas in Harrietella both sexes display a 1-segmented endopod and no sexual dimorphism in setation is discernible ( e.g. Vervoort 1950; Hicks 1989). The 2-segmented P3 endopod in the male, displaying a straight spinous outgrowth on the distal segment, is indicative of a relationship with Pseudonychocamptus , Pilifera , Inermiphonte , gen. n. (see below) and Marbefia , gen. n., however, Harrietella does not display the sexual dimorphism (on P3–P4 enp-1) that defines Lee and Huys’ (1999) PWPH -clade. Although this absence is conceivably secondary as a result of the loss of the inner seta on P3 enp- 1 in the female, confirmation of its membership of this clade will probably have to await the arrival of molecular sequence data. Harrietella is here tentatively regarded as the sistergroup of Inermiphonte , gen. n. (see below).
Although the mosaic distribution of unique plesiomorphies in single species [e.g. Heteronychocamptus exiguus ( Sars, 1905) with inner seta on P2 enp-1 ( cf. Sars 1905: Fig. 125); Laophonte danversae Hamond, 1969 with 5 elements on ♁ P5 exopod ( cf. Hicks 1982: Fig. 6 View Figure 6 ); P. carthyi with 3 inner setae on ♀ P3 enp-2 ( Figure 5C View Figure 5 )] hampers the analysis of the basal relationships within the PWPH -clade, a well defined group of five genera can be distinguished based on the sexual dimorphism of the P3 endopod. In males of Marbefia , gen. n., Pseudonychocamptus , Pilifera , Inermiphonte , gen. n. and Harrietella the ramus is 2-segmented (instead of 3-segmented), bearing a straight (rather than a sigmoid) apophysis on the outer margin of the distal endopod segment (Table 1). Within this lineage, Marbefia is considered to be the basal offshoot, the other four genera sharing a suite of synapomorphic reductions on the swimming legs: (1) P3–P4 exp-3 with 1 inner seta, (2) P2 enp-1 with 1 inner seta, and (3) P3 enp- 2 ♀ with at most 2 inner setae.



| R |
Departamento de Geologia, Universidad de Chile |
| MB |
Universidade de Lisboa, Museu Bocage |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Marbefia carthyi ( Hamond, 1968 )
| Huys, Rony & Lee, Wonchoel 2009 |
Pseudonychocamptus carthyi
| Hamond 1968 |