
Pugettia Dana, 1851
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3693.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:4E9BEE4F-4911-497C-BA86-DEC88DF5578E |
|
DOI |
https://doi.org/10.5281/zenodo.6146262 |
|
persistent identifier |
https://treatment.plazi.org/id/930FED1E-9704-FFCA-4183-FE58F0884C7B |
|
treatment provided by |
Plazi |
|
scientific name |
Pugettia Dana, 1851 |
| status |
|
Genus Pugettia Dana, 1851 View in CoL
Pugettia Dana, 1851: 268 (type species Pugettia gracilis Dana, 1851 , subsequent designation by Miers 1879).—Miers 1879: 650.—Stimpson 1907: 24.—Holmes 1900: 24.—Rathbun 1925: 167.—Schmitt 1921: 205.—Garth 1958: 186.—Sakai 1965: 72.—Griffin & Tranter 1986: 92.—Ng et al. 2008: 101.—Wicksten 2012: 209.
Mimulus Stimpson, 1860: 199 .—A. Milne-Edwards 1878: 144.—Holmes 1900: 23.—Rathbun 1904: 173.—Rathbun 1925: 182.—Schmitt 1921: 204.—Garth 1958: 183.—Ng et al. 2008: 101.—Wicksten 2012: 208.
Type locality Puget Sound, Washington.
Species included
Pugettia elongata Yokoya, 1933
Pugettia foliata (Stimpson, 1860) Pugettia gracilis Dana, 1851 (type species) Pugettia hubbsi Garth, 1958
Pugettia incisa ( De Haan, 1839)
Pugettia intermedia Sakai, 1938
Pugettia kagoshimensis Rathbun, 1932 Pugettia leytensis Rathbun, 1916
Pugettia marissinica Takeda & Miyake, 1972 Pugettia mindanaoensis Rathbun, 1916 Pugettia minor Ortmann, 1893
Pugettia nipponensis Rathbun, 1932 Pugettia ogasawaraensis Komatsu, 2011 Pugettia producta (Randall, 1840) Pugettia quadridens ( De Haan, 1839) Pugettia pellucens Rathbun, 1932 Pugettia richii Dana, 1851
Pugettia similis Rathbun, 1932
Pugettia tasmanensis Richer de Forges, 1993 Pugettia venetiae Rathbun, 1924
Remarks. In his Latin description (translated by Holmes 1900), Stimpson (1860) defined the genus as: “ Carapace flattened, more or less pentagonal; antero-lateral margin laminate and cut by a narrow fissure into two closely approximate lobes. Rostrum short, bifid, and horizontal. Orbits incomplete below, but furnished above with a preorbital and postorbital spine. Eyes not concealed when retracted. Merus of the external maxillipeds short, the external angle obtuse, the internal angle incised; outer margin of the exognath dilated. Hand of the chelipeds much compressed and sublaminate. The propodi of the ambulatory legs have a setose tooth near the middle of the inferior margin. First pair of ambulatory legs exceeding the others.”
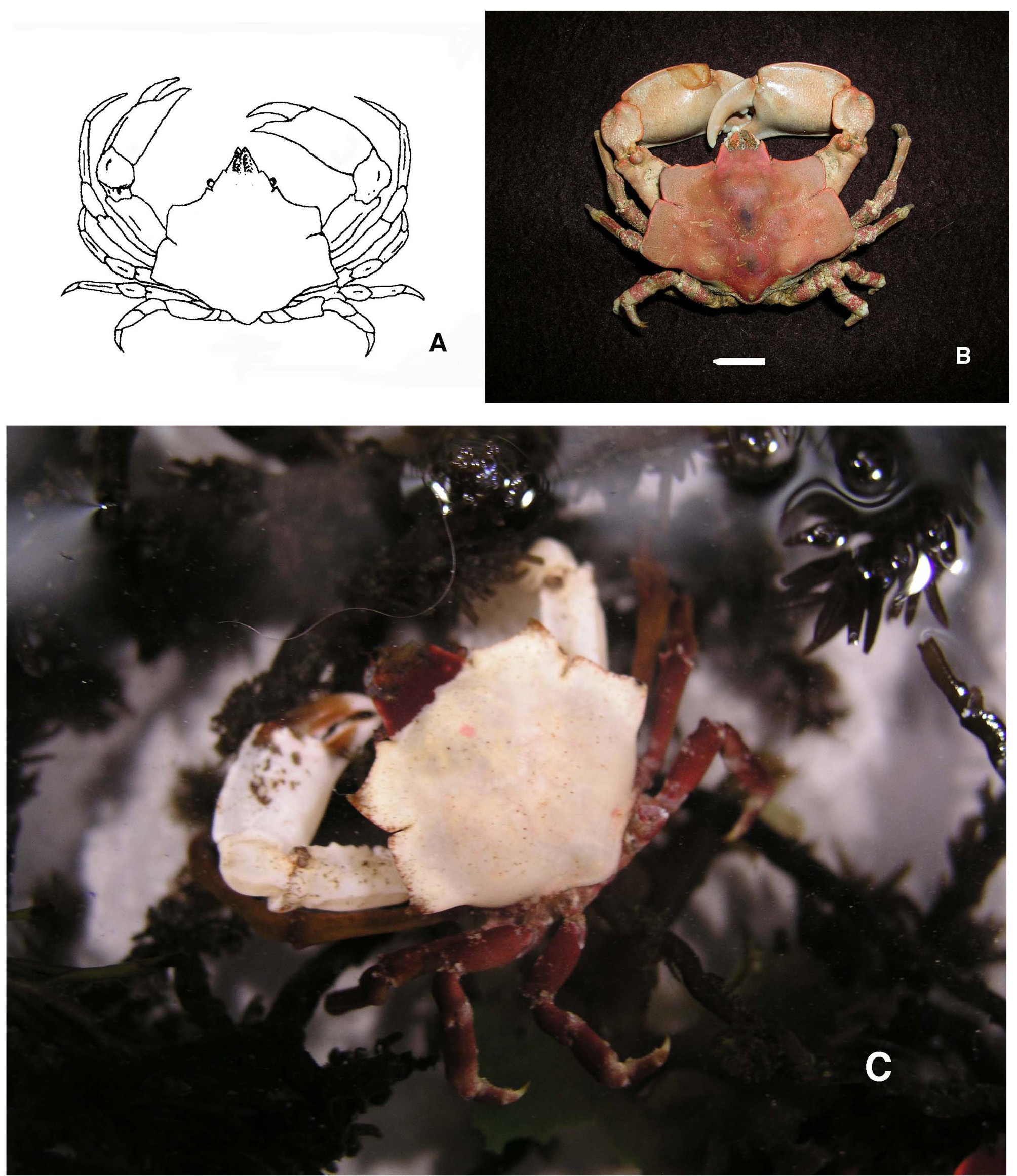
Stimpson stated "this genus is near Epialtus , with lamellar expansions of the sides of the carapax as in some Hueniae ". The narrow chelipeds shown in the original illustration demonstrate that this individual was a female (Stimpson 1860: pl.1A). The type locality was “taken from the stomachs of percoid fishes, (“Cabesones”) caught off Monterey, California, by A.S. Taylor”. Cabezon is the common name of Scorpaenichthys marmoratus (Ayres, 1854) , family Cottidae , a crab-eating fish. Stimpson made no mention of or made comparison to species of Pugettia . Stimpson’s specimens were housed in the Chicago Academy of Sciences, which was destroyed in a fire in 1871. The type specimen almost surely was destroyed. Garth (1958) reported that a “male cotype without chelipeds or legs” existed in the Museum of Comparative Zoology, Harvard University, but no syntypes of M. foliatus have been found in the British Museum (Natural History) (Evans 1967).
A. Milne-Edwards (1867) described a second species, Mimulus acutifrons , which supposedly could be distinguished from M. foliatus by the “frontal teeth” (rostral horns), which are “short and hardly separated from the median line, and by the supra-orbital angles not reaching as far” (translated from the French). There were no illustrations and there was only a single specimen (sex not stated), and no information on the type locality was given except that the specimen had been part of the collections of “Baron de Lafresnaye”. The type material, which seems to have been deposited in the Muséum National d’Histoire Naturelle, Paris, is apparently lost (D. Guinot, pers. comm.) Garth (1958) questionably placed M. acutifrons in synonymy with M. foliatus . The length of the rostral horns is variable in specimens of M. foliatus , and the supra-orbital angles can vary with age and sex. It is impossible without the examination of the holotype to know for certain if Milne-Edwards’ specimen could be assigned to Mimulus or a related genus, or in which localities one might look for a similar specimen.
Newcombe (1893) contacted M.J. Rathbun regarding a specimen collected in Clayoquot Sound, British Columbia. She stated that she was “unable to separate the genus from Pugettia ”. Newcombe (1893) recorded the species as Pugettia (Mimulus) foliata Stimpson. Rathbun (1894:72) wrote "there seems to be no good reason for placing this ( Mimulus foliatus ) in a genus distinct from Pugettia ," and referred to the species as Pugettia foliata . The "antennae have the basal joint as in that genus" ( Pugettia ) and the flagellum is flattened and exposed at the side of the rostrum, the "carapace is wider than in other species of Pugettia " and the chelipeds present showed "nothing distinctive". The maxillipeds, abdomen, sternum, and ambulatory legs were considered as "almost exactly" as in Pugettia gracilis Dana, 1851 .
Holmes (1900: 23) translated Stimpson's original Latin description of Mimulus but made no comparisons to species of Pugettia . Without explanation, he cited Pugettia foliata , as given by Rathbun (1894), as a junior synonym of M. foliatus . Rathbun (1904: 173) referred it as M. foliatus . Schmitt (1921), Rathbun (1925), and Garth (1958) considered Mimulus a valid genus, containing only one species, M. foliatus . Garth’s work provided a brief re-description of the species along with illustrations, and Garth (1958) and Wicksten (2012) provided extensive synonymies.
Schmitt (1921), Rathbun (1925), and Garth (1958) distinguished species of Pugettia from Mimulus on the basis of the lateral margins of the carapace being markedly flattened or produced. Schmitt (1921) stated that the upper surface of the carapace of Pugettia bore spines or tubercles instead of being smooth, and assigned P. producta Randall, 1839 , to Epialtus H. Milne-Edwards, 1834 . The dorsal surface of the carapace of P. producta is smooth. Schmitt's description of M. foliatus noted that it possessed two median "obsolescent" tubercles as well as a tubercle on the posterior branchial region, and thus contradicted the generic distinction from Pugettia given in his key.
Garth (1958: pls. L, O, P, Q) figured the right first pleopods of male epialtid crabs (as subfamilies Acanthonychinae and Pisinae of the Majidae ), including M. foliatus , P. producta , P. richii , four other Pugettia species, and Talipeus nuttallii . In both M. foliatus and the species of Pugettia , this pleopod bears a flared apex with a pointed tip and one or two opposing grooved projections of equal or slightly shorter length. Both M. foliatus and the species of Pugettia bear some form of median protuberance: a spinulous lobe, "tongue," rounded lobule, or two smaller lobes. In contrast, the first pleopods of T. nuttallii do not have the median protuberance, and the lateral projections are not curved, as in Pugettia . The male pleopods of other species of Epialtidae vary, ending in a curved, spiny, or flattened apex, two concave points, small and blunt lobes or a complicated structure of depressed areas and protrusions. The shapes of the distal areas of the male first pleopods of M. foliatus , P. producta P. richii , and T. nuttallii are in close agreement with those figured by Garth. Griffin & Tranter (1986: fig. 28) figured the male first pleopods of the western Pacific P. incisa , P. intermedia , P. marissinica , and P. quadridens . Komatsu (2011), in the original description of P. ogasawarensis , and Richer de Forges (1993), in the original description of P. tasmanensis , also figured the first pleopods. The pleopods of all of the western Pacific and Tasman Sea species have similar apices, only varying in fine details of the angle, length, and curvature of the lobes and tubercles. The structure of the first pleopod of male M. foliatus falls within the range of species variation of the genus Pugettia .
Hultgren & Stachowicz (2008a) studied the habitat distinctions between P. producta , P. richii , and M. foliatus . These three species can be found in intertidal areas along the coast of California, but have distinct habitats: P. producta is usually found among large kelps, ( Macrocystis or Egregia spp.), P. richii among red algae (Rhodophyta), and M. foliatus typically in the lowest intertidal zone into shallow subtidal regions, on rocks, among algae, or in kelp holdfasts ( Fig. 1 View FIGURE 1 C). The relatively smooth carapaces of P. producta and M. foliatus may be related to climbing on algae or hiding among kelp holdfasts. Of the three species, only P. richii decorates by attaching algae or hydroids to its rostrum and the sides of the carapace. The other two species may attach a small piece of alga to the rostrum but do not cover the body from dorsal view. The three species share similar ranges: P. producta from the Queen Charlotte Is., Canada to Point Asunción, Baja California, Mexico; P. richii from Prince of Wales I., Alaska to Asunción Bay, Baja California; and M. foliatus from Unalaska, Alaska to San Diego, California, although it is uncommon south of Point Conception, California. A report of M. foliatus from Mazatlán, western Mexico is "questionable" (Garth 1958).
Hultgren & Stachowicz (2008b) used a genetic analysis to study the relationships among majoid crabs. The study included members of the Epialtidae , including six species of Pugettia , M. foliatus , and Taliepus nuttallii . It was found that M. foliatus was as closely related to other species of Pugettia as species of Pugettia were to each other. As pointed out by Rathbun (1894: 72), the closest relative to M. foliatus seems to be P. gracilis . Hultgren & Stachowicz (2008b) suggested, but did not confirm, that Mimulus should be considered to be a junior synonym of Pugettia .
Stimpson (1860) and subsequent authors gave great importance to the lateral flattened expansions of M. foliatus , as seen in the "Hueniae", as a difference with other majoid crabs. The first author examined specimens of Huenia heraldica ( De Haan, 1837) (Epialtidae) in the collections of the Bishop Museum, Honolulu, Hawaii. In H. heraldica , the shape of the lateral expansions of the carapace is sexually dimorphic, with those of the female being less sharply defined than those of them male. These lateral expansions occur in other epialtoid crabs, such as Epialtoides hiltoni (Rathbun, 1894) .
Large, well-separated teeth along the lateral margin of the carapace in most species of Pugettia , but in P. gracilis , the anterolateral projections of the carapace have been described as "wing-like" (Garth 1958). Hultgren & Stachowiz (2008a) related the surface of the carapace to habitat, suggesting that crabs that climb among algae or hide under rocks might be more likely to have a smooth carapace than those living among dense algae. The function of the “wing-like expansions” of the carapace remains uncertain, but combined with a color similar to the algae on or among which they live, may help to conceal the crab in the natural habitat.
As in M. foliatus , adult male Pugettia frequently have larger chelae than do mature females, often with a gape between the fingers, and a pronounced ridge along the carpus and the upper margin of the propodus ( Fig. 1 View FIGURE 1 B). These features are prominent in P. richii , P. dalli , and P. gracilis from the eastern Pacific; somewhat less so in P. producta and P. hubbsi ; and in the Japanese species P. minor , P. incisa , P. quadridens , P. nipponensis , and P. sagamiensis (Sakai 1965: pls. 31–33).
Species of Pugettia , with the exception of P. tasmanensis , are confined to the western and northern Pacific Ocean, ranging from the Philippines northward along the Asian coast, across the Aleutian Islands south to Baja California, Mexico. Thirteen of the 21 species inhabit the area from the Philippines to Russia, and seven are found in the eastern Pacific. P. tasmanensis is the only species reported from the southern hemisphere. In California, these are called “kelp crabs”, an appropriate name because many of them live among algae.
Based on external morphology, structure of the male first pleopods, sexual dimorphism, habitat, range and new genetic data, we find no reason to separate Mimulus foliatus from the known species of Pugettia . We therefore place Mimulus Stimpson, 1860 in synonymy with Pugettia Dana, 1851 . The revised name of the foliose kelp crab is hereby changed to Pugettia foliata (Stimpson, 1860) .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
SubFamily |
Epialtinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
SubFamily |
Epialtinae |