
Taenionema, Banks, 1905
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4767.3.4 |
|
publication LSID |
urn:lsid:zoobank.org:pub:D8ACE0A9-A837-4138-9642-E6C0086DB017 |
|
DOI |
https://doi.org/10.5281/zenodo.3797054 |
|
persistent identifier |
https://treatment.plazi.org/id/966587F2-FF97-FFA4-12D4-3B95ABBFFEDB |
|
treatment provided by |
Plazi |
|
scientific name |
Taenionema |
| status |
|
Figs. 1–11 View FIGURE 1 View FIGURE 2 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 .
Material examined. One male larva, two female larvae, China: Zhejiang Province, 15 km SE. Quzhou City, Mt. Lankeshan, stream in scenic area ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), 28° 52′ 42.63″ N, 118° 55′ 22.45″ E, 250 m, March 14–15, 2020, leg. Zhi-Teng Chen, Xiao-Han Ye (ICJUST).
Description. Mature larva. Body length ca. 7 mm (n = 1). General color brown, head and abdomen darker than thorax (Fig. 3).
FIGURE 3. Taenionema sp., male larva, A. larval habitus, dorsal view; B. larval habitus, ventral view.
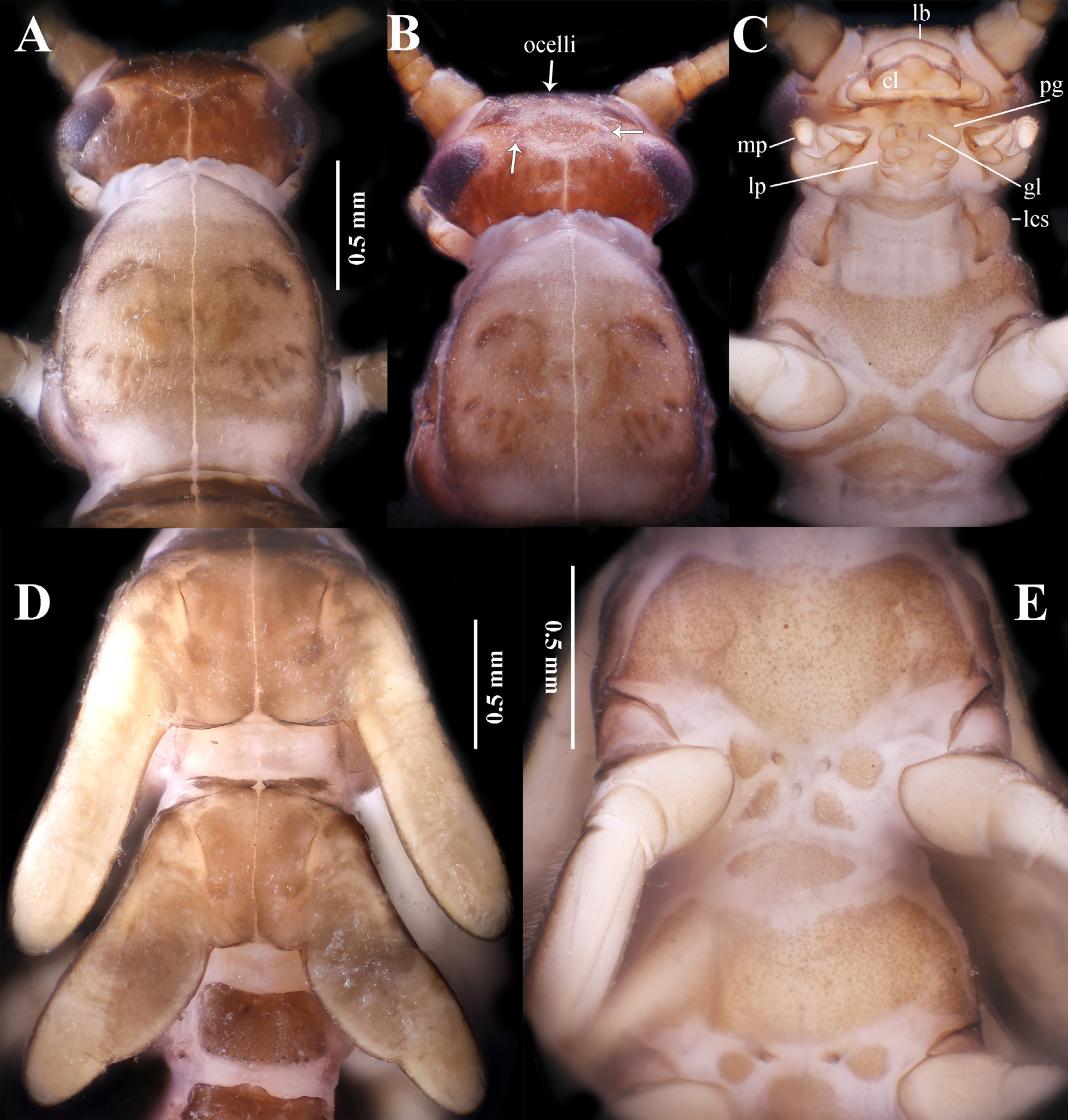
Head. Head ( Fig. 4 View FIGURE 4 A–C) oval and covered with pale short hairs, frontal area with a U-shaped brown pigmented area; labrum broad and short; clypeus broad, with three lobes and a truncate anterior margin. Three ocelli pale, subequal in size, with brown margins; each posterior ocellus with a semicircular anterior stigma; compound eyes dark and glabrous; area between each compound eye and posterior ocellus pale. Occiput scattered with obscure brown rugosities. Antennae slender, subequal in length to abdomen, basal half pale, apical half light gray-brown, each segment with very short cylindrical apical spines ( Fig. 5 View FIGURE 5 ).
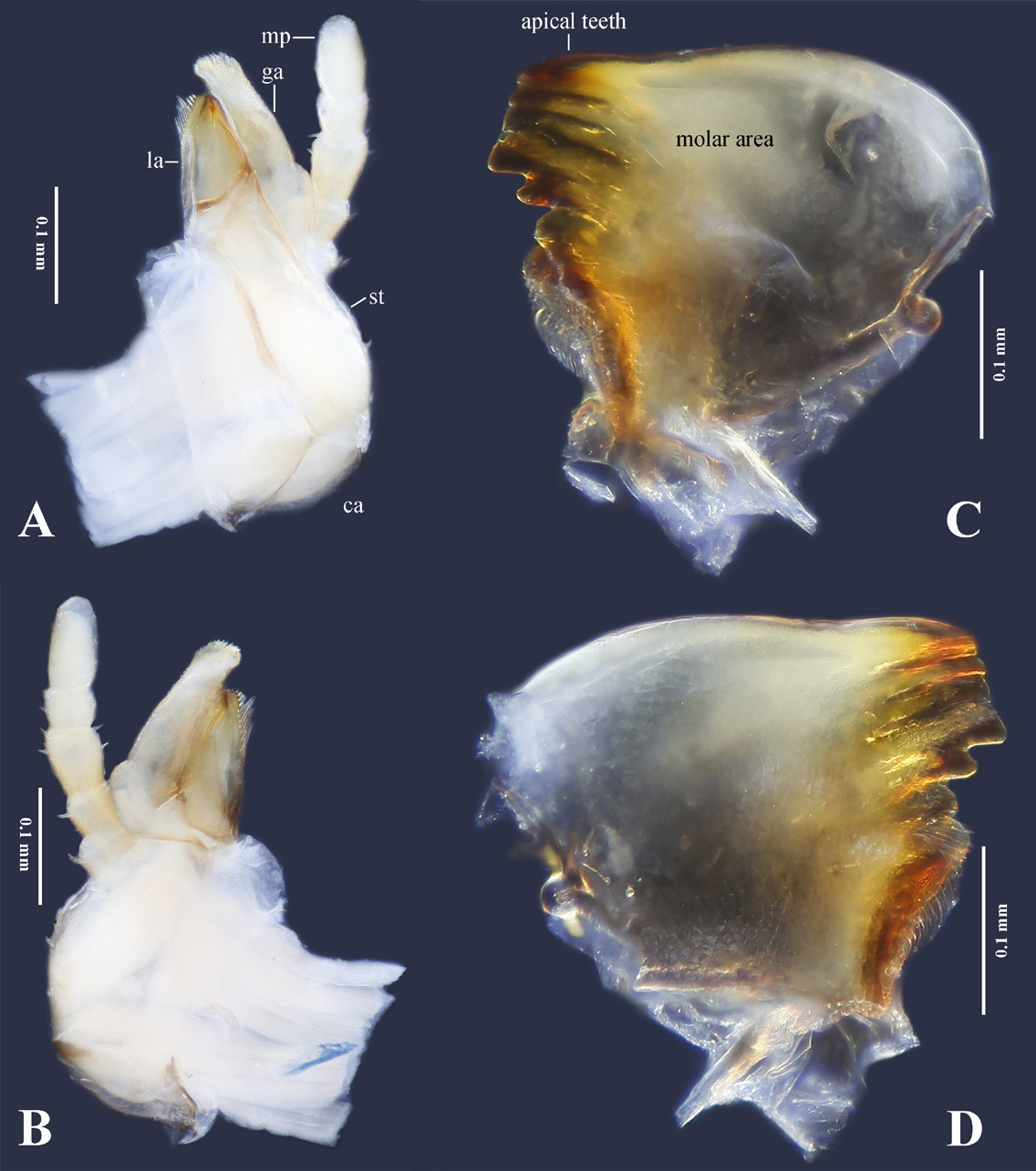
Mouthparts typical euholognathan, glossa and paraglossa knob-shaped, equally short ( Figs. 4C View FIGURE 4 , 6 View FIGURE 6 ); labial palp very short, three-segmented with basal segment being shortest; maxillary palp five segmented; stipes of maxilla long cylindrical, basal half fringed with sparse short bristles, basally with a patch of short bristles ( Fig. 6 View FIGURE 6 A–B); galea lateral apex densely with short spines, medial margin without spines, glabrous; lacinia semicircular and sclerotized with short, stout apical teeth, dorsal comb with about 10 long bristles. Mandible consists of five major unserrated, sclerotized apical teeth, posterior to these are two shorter teeth, a submarginal bunch of about eight long spines and a marginal comb of about 10 long spines; molar area unmodified ( Fig. 6 View FIGURE 6 C–D). Cervical membranes long with a conspicuous median sclerite.
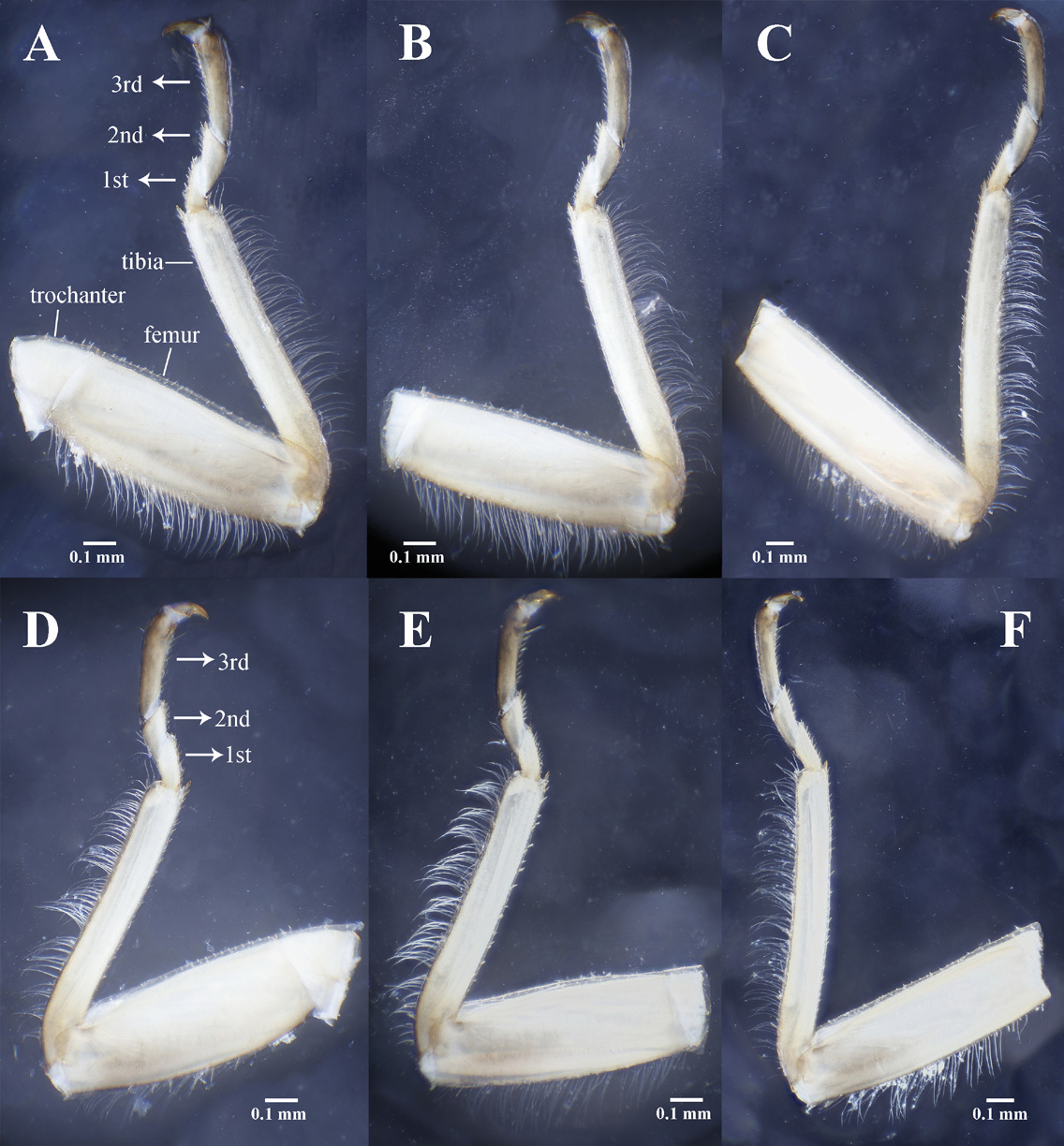
Thorax. Pronotum disc near oval with posterior margin truncate ( Fig. 4A, B View FIGURE 4 ), mostly pale brown with dark rugosities, densely covered by pale spinules. Meso- and metanota near glabrous, with a few pale hairs at anterior areas ( Fig. 4 View FIGURE 4 D–E); wing pads of mesonotum rodlike, slightly constricted at mid-length, apical half covered with sparse dark spines; wing pads of metanotum notched along posterior margin, apical half covered with sparse dark spines; ventral thoracic sclerites clearly visible, mesothoracic Y-arms and stem forming three deep pits. Coxae membranous and glabrous dorsally, ventrally and laterally spinulose and weakly sclerotized; trochanters ventrally spinulose. Femora dorsally covered with dense short spines except for a glabrous longitudinal stripe ( Fig. 7 View FIGURE 7 A–C), median line with several larger spines, ventral aspect mostly glabrous except for several apical spines ( Fig. 7 View FIGURE 7 D–E), outer margin with dense swimming hairs which shortened towards each end of femora, inner margin with two glabrous ridges. Tibiae dorsally covered with dense spinules, ventrally with an irregular row of spines, outer margin with dense swimming hairs; two large tibial spurs present ventrally ( Fig. 7 View FIGURE 7 ). Tarsal segments darkly sclerotized, mostly glabrous, inner margin with dense spines, outer margins with sparse short swimming hairs ( Fig. 7 View FIGURE 7 ).
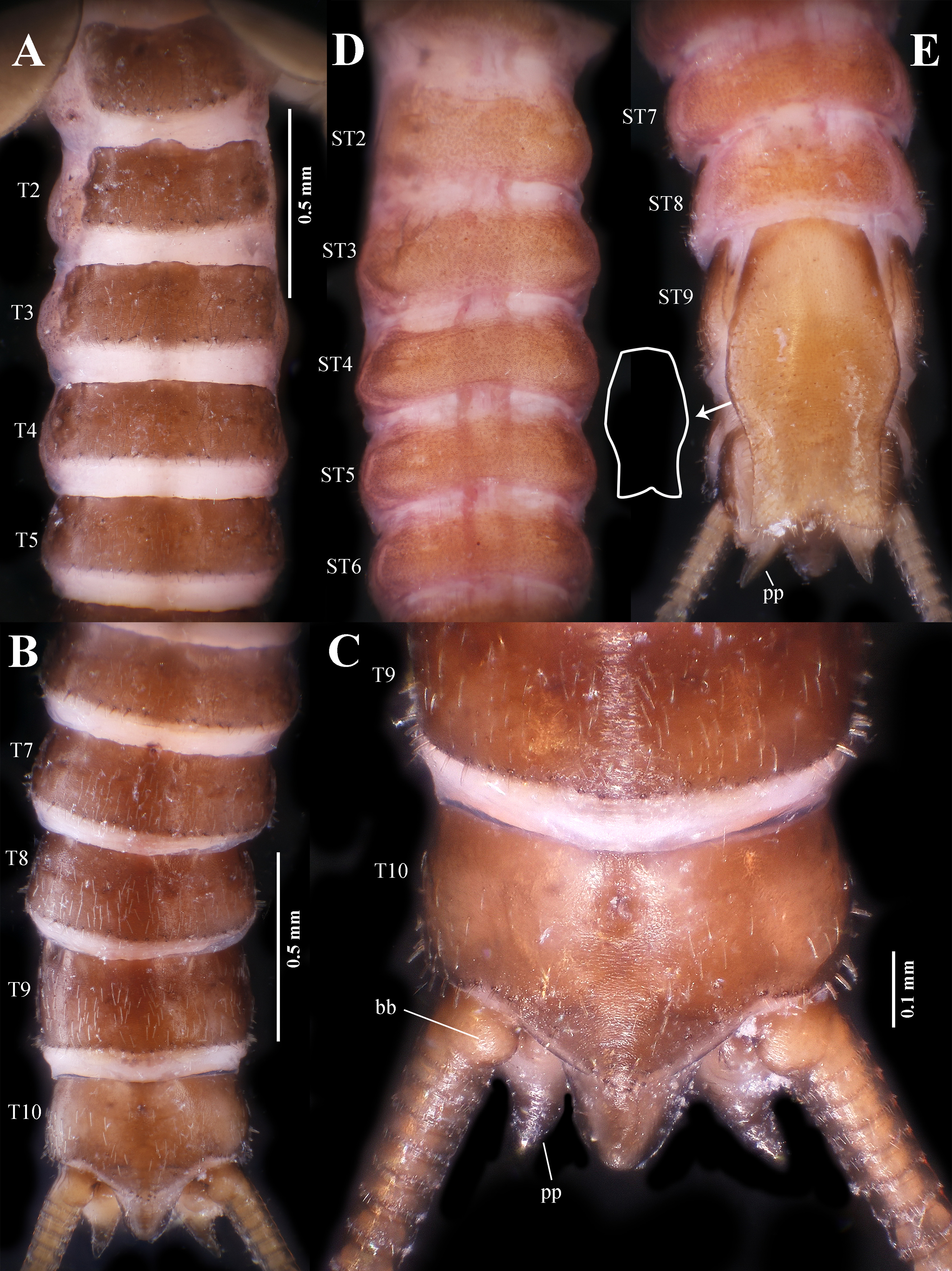
Abdomen. Posterior half of abdominal terga 1–6 covered with short black hairs (Figs. 3A, 8A–B); terga 2–5 with medially projected anterior margins; terga 7–9 covered with sparse pale hairs; posterior margins of terga 1–9 fringed with dark, stout spines. Tergum 10 generally glabrous ( Fig. 8 View FIGURE 8 B–C), concave medially, posterolateral margins fringed with pale, stout spines; posterior margin of tergum 10 strongly projected backwards, forming a membranous process. Sterna 1–8 spinulose (Figs. 3B, 8D–E). Median area of sternum 9 with a large post-genital plate exceeding basal cercal segment ( Fig. 8E View FIGURE 8 ); the plate with a trapezoidal base which is evenly covered with short hairs, apical half constricted and covered with much longer pale spines, apical ¼ with parallel lateral sides and terminated with a subtriangular shallow notch. Paraprocts glabrous and drop-shaped, apices abruptly constricted into sharp projections ( Fig. 8C View FIGURE 8 ). Cerci pale, generally glabrous, subequal in length to abdomen; basal cercal segment much longer than other basal segments (at least 2–10), enclosing rounded basal cercal bulb of male adult; each segment with very short cylindrical apical spines (Figs. 3, 9).
Female larva. Body length ca. 10 mm (n = 2) ( Fig. 10 View FIGURE 10 ). Generally similar to male. Abdominal terga 1–9 each with a transverse row of dark pits ( Figs. 10A View FIGURE 10 , 11A View FIGURE 11 ). Tergum 10 covered by dark spines, medially with an anchorshaped pale area, posterior margin extended backwards but not exceeding 1.5X of the length of tergum 10 ( Fig. 11A View FIGURE 11 ); a rounded membranous process present beneath tergum 10. Ventral thoracic sclerites invisible, instead with granular and spinulose areas ( Fig. 10B View FIGURE 10 ). Sternum 8 mostly glabrous, medially with a U-shaped sclerite, covering the ovipore of female adult ( Fig. 11 View FIGURE 11 B–C). Post-genital plate not exceeding basal cercal segments, its base being mostly glabrous and constricted in the apical; the plate with a sub-trapezoidal base that is mostly glabrous, its apical half constricted and covered with pale, long spines, the apex rounded or truncate ( Fig. 11 View FIGURE 11 B–C). Paraprocts glabrous, with rounded apices ( Fig. 11 View FIGURE 11 B–C).
Distribution. Southeastern China (western Zhejiang Province).
Diagnosis. The larvae are assigned to Taeniopterygidae by the presence of a combination of characters: the subequal length of the glossae and paraglossae, the divergent wing pads, and the second tarsal segment being as long or longer than the first ( Stewart & Stark 2002)). The absence of coxal gills supports the larvae being included in the subfamily Brachypterainae ( Ricker & Ross 1975) . The combination of characters including brown body color with darker brown mottling on head and thorax and lack of silky cercal fringe hairs support the larvae being a species of Taenionema ( Stewart & Stark 2002, Stewart 2009). Kyphopteryx and Mesyatsia , the two brachypteraine genera known from China, have absent or reduced anterior ocellus that could also be revealed by mature larvae (see fig. 7 in Teslenko & Zhiltzova 1992). The presence of three well-developed ocelli in Taenionema sp. ( Fig. 4B View FIGURE 4 ) has excluded its recognition as Kyphopteryx or Mesyatsia . Adult basal cercal bulbs and detached cerci are already clearly visible inside exoskeleton of the mature male larva of Taenionema sp. ( Fig. 8 View FIGURE 8 B–C), whereas abdominal terga 2–3 remain flat and unmodified ( Fig. 8A View FIGURE 8 ), covering no toothed processes as diagnostic in Kyphopteryx . In Mesyatsia , despite the reduced anterior ocellus, the larvae of M. makartchenkoi Teslenko & Zhiltzova, 1992 have shown very long silky cercal hairs on each segment ( Teslenko & Zhiltzova 1992, 2009), which is different from the larvae described herein ( Fig. 9 View FIGURE 9 ). Male larva of Taenionema sp. appear different from its Nearctic congeners by the unique, emarginate post-genital plate (see fig. 365 in Judson & Nelson 2012) and hook-shaped paraprocts ( Stewart 2009). When compared with the eastern Palearctic T. japonicum , male larva of Taenionema sp. also seems unique by the mandible having more apical teeth and two additional basal combs of long spines on mandible (see fig. 1671 in Teslenko & Zhiltzova 2009), different head pattern and distinctly elongated pronotum (see fig. 371 in Judson & Nelson 2012). However, accurate species determination of Taenionema sp. is tentative until the association of adults and larvae are confirmed.
Remarks. Larvae of Taenionema sp. were collected from leaf packs and woody debris along banks of a large stream in slower current speeds ( Fig. 1 View FIGURE 1 C–D). Two days of sampling revealed this species to be in low density. Larvae of Nemouridae , Leuctridae and Perlidae were also present. We attempted to rear the larvae to obtain adults, however, after three days the larvae failed to emerge and were preserved before returning to the laboratory. Regardless, the discovery of Taenionema in China, especially from a low-elevation coastal stream is unquestionable. Adult morphology will be supplemented when material is obtained.
Acknowledgments
We are grateful to Prof. Yu-Zhou Du and Yangzhou University for teaching us the taxonomy of the Plecoptera . We are also grateful to Drs. Boris Kondratieff (Colorado State University) and R. Edward DeWalt (University of Illinois) for helpful comments and manuscript improvement. This work is funded by the Start-up Funding of Jiangsu University of Science and Technology (1182931901).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |