
Careproctus, KrOyer, 1862
|
publication ID |
https://doi.org/10.12782/specdiv.29.111 |
|
publication LSID |
lsid:zoobank.org:pub:C8E2C4A5-D776-4995-B139-BC2C56B5D133 |
|
persistent identifier |
https://treatment.plazi.org/id/9816756E-FFAC-FFB3-FC43-FB7FFB01F862 |
|
treatment provided by |
Felipe |
|
scientific name |
Careproctus |
| status |
|
Careproctus View in CoL View at ENA io sp. nov.
[New standard Japanese name: Mini-kon’nyakuuo] ( Figs 1 View Fig , 2 View Fig ; Table 1)
Holotype. FAKU 148130 View Materials , 37.3 mm SL ( 44.2 mm TL), female, off Iwaki , Fukushima, Japan, 36.9359°N, 141.5477°E, 419 m depth, 13 November 2020, otter trawl, R / V Wakataka-maru, coll. R. Misawa and E. Morikawa. GoogleMaps
Paratypes. Two specimens: FAKU 150259 View Materials , 42.8 mm SL ( 51.3 mm TL), male, off Ofunato , Iwate, Japan, 39.0360°N, 142.1736°E, 465m depth, 19 June 2023, beam trawl, R / V Kaiyo-maru No GoogleMaps . 6, coll . E GoogleMaps . Morikawa ; FAKU 150605 View Materials (stained and dissected, used only for osteological description), 33.3 mm SL (TL unknown), female, same data as FAKU 150259 GoogleMaps .
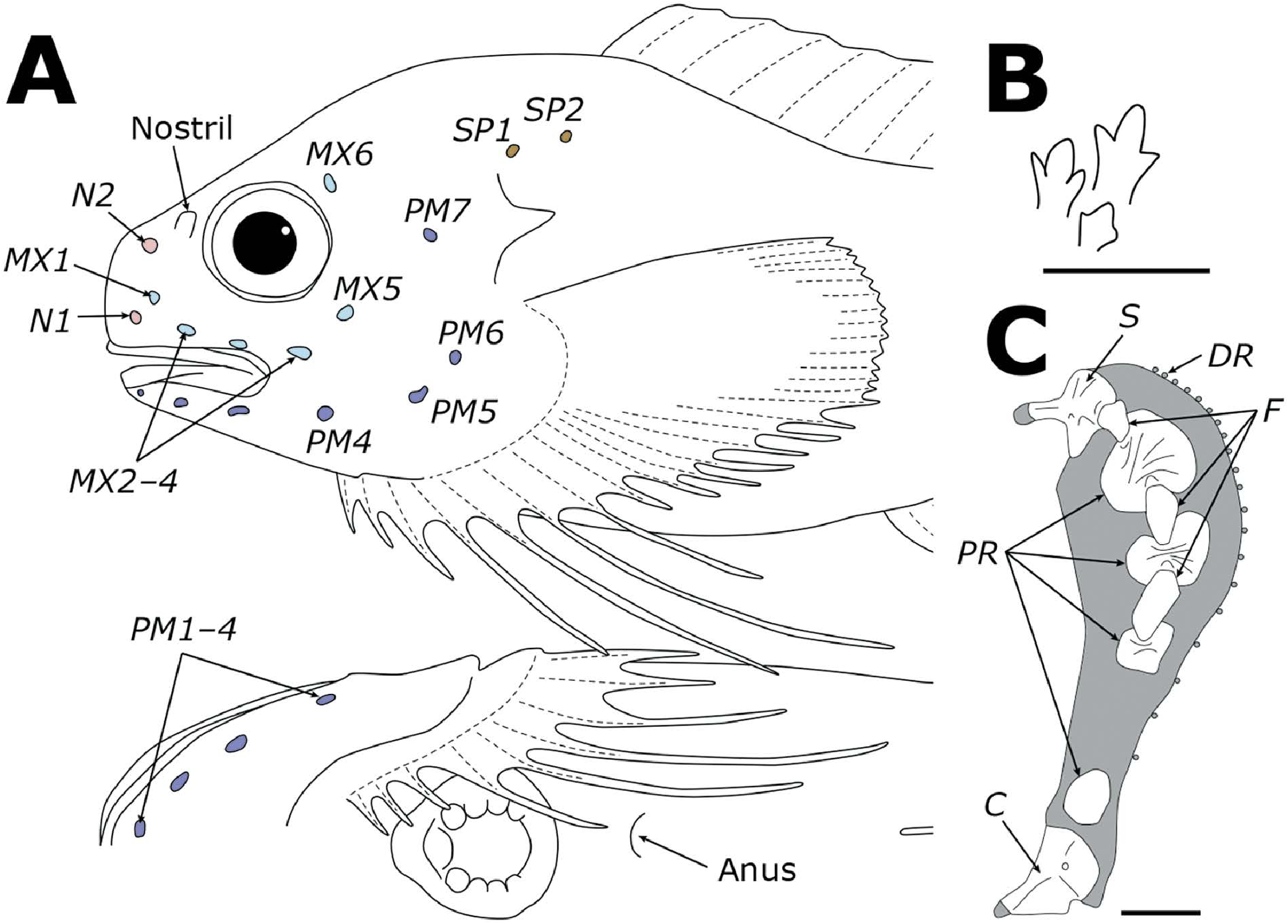
Diagnosis. A species of Careproctus with the following combination of characters: vertebrae 40–42; dorsal-fin rays 36 or 37; anal-fin rays 30; pectoral fin with 28 or 29 rays, deeply notched, lower lobe reaching to anal-fin origin; large pelvic disk 34.2%–34.5% HL (10.3%–10.9% SL); teeth strongly trilobed on both jaws, inner teeth weakly trilobed or shouldered; cephalic pore pattern 2-6-7-2, chin pores paired; gill slit above pectoral-fin base; body bright red, non-variegated in life.
Description. Counts and measurements shown in Table 1. Data for holotype given first, followed by paratype data in parentheses, if different.
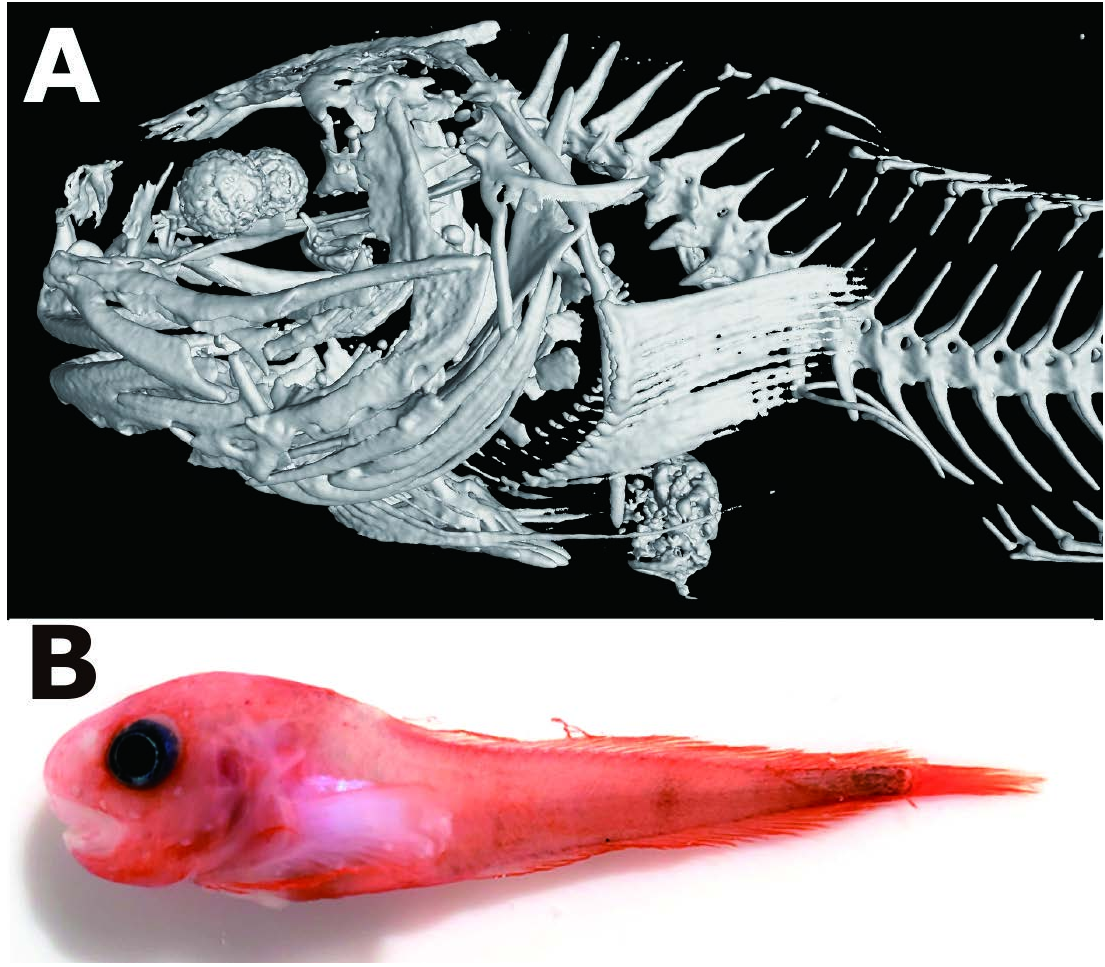
Body short, robust, subcylindrical anteriorly, slightly compressed posteriorly; dorsal profile humpbacked, gradually sloping posteriorly, deepest at nape ( Fig. 1A View Fig ). Head robust, rounded. Eye large, upper margin slightly below dorsal contour. Interorbital space almost flat. Skin relatively thin, without subcutaneous gelatinous layer; prickles absent. Head robust, large, dorsal profile strongly sloping from nape to snout. Interorbital region flat, smooth. Snout blunt, protruding slightly beyond tip of upper jaw. Mouth subterminal; maxilla extending to mid orbit; oral cleft extending to anterior margin of orbit. Premaxillary teeth strongly trilobed, in 8 oblique rows forming broad bands; inner teeth becoming larger, weakly trilobed or shouldered ( Fig. 1B View Fig ). Mandibular teeth trilobed in 8 oblique rows; inner teeth becoming larger. Diastema absent at symphysis of upper and lower jaws. Orbit round, moderately large; pupil large, rounded. Nostril single, with short tube at level of mid orbit. Pore size of cephalic lateralis moderate: nasal pores 2, maxillary pores 6, preoperculo-mandibular pores 7, suprabranchial pores 2; cephalic pore pattern 2-6-7-2 (damaged in paratypes) ( Fig. 1A View Fig ). Chin pores paired in separate pits. Coronal pore absent. Gill slit small, upper margin level with dorsal rim of orbit, extending ventrally to just above pectoral-fin base. Opercular flap angular, pointed posterodorsally.
Vertebrae 42 (40), precaudal 10 and caudal 32 (30). Dorsal fin without lobes, rays 36 (37), tip of rays slightly exserted. Anteriormost dorsal-fin pterygiophore inserted between neural spines 3 and 4 (or 2 and 3), bearing single short ray ( Fig. 2A View Fig ). Membrane of posterior dorsal-fin rays continuous with caudal fin. Anal-fin rays 30. Anal-fin origin below vertebrae 12. Membrane of posterior anal-fin rays continuous with caudal fin. Caudal fin slightly rounded. Principal caudal-fin rays 10 (11); dorsal procurrent rays 2 (3), ventral procurrent rays 2 (1). Hypurals and parhypural fused into single plate; its upper and lower portions separated by narrow slit. Pleural ribs in 2 pairs, on abdominal vertebrae 9 and 10 (8 and 9).
Pectoral fin deeply notched, with 28 (29) rays, just reaching to level of anal fin origin; upper lobe with 20 rays; lower lobe moderately elongate, with 8 (9) rays, 6th ray from ventralmost longest, just reaching to anal-fin origin. Tip of pectoral-fin rays free of membrane, lower rays more strongly exserted; rays in notch slightly more widely spaced than rays of lobes. Uppermost pectoral-fin base level with ventral rim of orbit. Lowermost pectoral-fin base below posterior rim of orbit. Proximal pectoral radials 4 (3 + 1), curved triangular (based on FAKU 150605; Fig. 1C View Fig ). Upper and lower part of radials 1 and 2 notched; upper part of radial 3 deeply notched; radial 4 without notch. Interradial fenestrae 3, extending between scapula and proximal radials 1–3; fenestra 1 blunt triangular, fenestrae 2 and 3 elliptic. Scapula broadly Y-shaped, with strong helve, extending closely to uppermost proximal radial. Coracoid pear-shaped, with thin lamella. Pelvic disk large, round; length slightly greater than width. Anus posterior to gill slit, slightly closer to pelvic disk than to anal-fin origin. Pyloric caeca 9 (8).
Coloration ( Fig. 2B View Fig ). In life, head, body, and fins bright red; snout, lips, and posterior part of pectoral-fin base pale pink; caudal peduncle somewhat darker; pelvic disk pale pink; eye black. After preservation, head, body, and fins white; caudal peduncle somewhat darker; eye black; stomach and peritoneum white.
Reproduction. Ovary of holotype pouch-like, translucent whitish, with yolked eggs ( 0.50 mm maximum diameter). Testes of paratype ( FAKU 150605 View Materials ) enlarged, color creamy-white .
Distribution. Known only from the Pacific coast of northern Honshu Is., Japan (off Fukushima and Iwate) at depths of 419– 465 m.
Etymology. The specific name is a noun in apposition after Io, a Greek mythological character who wandered the world without rest, due to the unsettled phylogenetic position of the new species (see under Remarks). The standard Japanese name “Mini” reflects its small body size.
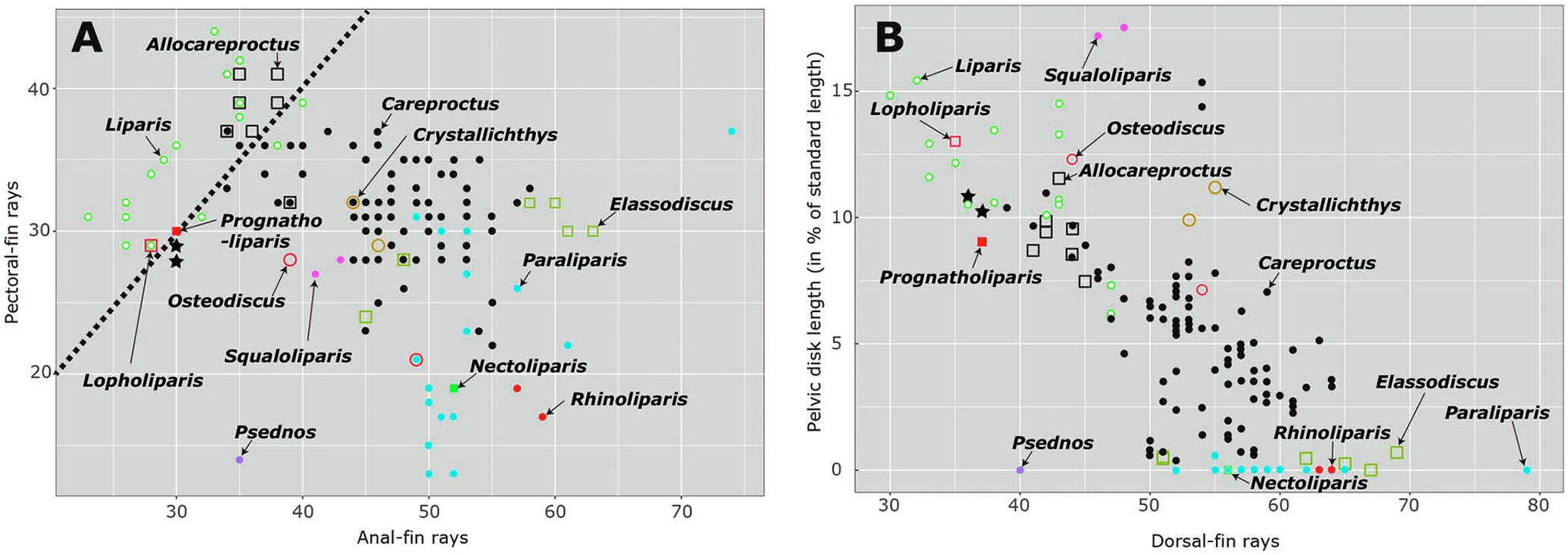
Remarks. The new species is characterized by similar numbers of pectoral- and anal-fin rays, reminiscent of Liparis , Allocareproctus Pitruk and Fedorov, 1993 , Prognatholiparis Orr and Busby, 2001 , and Lopholiparis Orr, 2004 ( Fig. 3A View Fig ). In addition, the pelvic disk size of the new species is relatively large for species of Careproctus , instead being similar to that of Liparis , Allocareproctus , and Prognatholiparis . Although species with a larger pelvic disk generally have a smaller number of dorsal-fin rays ( Fig. 3B View Fig ), plots of these two characters for the new species were not typical for Careproctus . However, C. io has a single nostril (vs. two nostrils in Liparis ), no pseudobranchs (vs. pseudobranchs present in Liparis ), a subterminal mouth (vs. lower jaw projecting in Prognatholiparis ), no papillae on rims of the cephalic lateralis pores (vs. papillae present in Allocareproctus ), and no enlarged cephalic bones (vs. enlarged cephalic bones present in Lopholiparis ), thereby clearly distinguishing the former from the latter genera ( Figs 1A View Fig , 2A View Fig ) ( Orr and Busby 2001, 2006; Orr 2004; this study).
In contrast, a BLAST search of the COI sequences (652 bp) of the new species showed them to be most similar to those of Pseudoliparis swirei Gerringer and Linley, 2017 (92.6%; INSDC accession numbers, KY659181–KY659184) and Notoliparis kermadecensis (Nielsen, 1964) (92.6%; KY659176, KY659179, KY659180). However, both Pseudoliparis Andriashev, 1955 and Notoliparis Andriashev, 1975 are characterized by a coronal pore ( Andriashev 1975; Andriashev and Pitruk 1993), clearly differing from the new species which lacks coronal pores. Neighbor-joining (NJ) and maximum likelihood (ML) trees reconstructed herein using the COI sequence of the new species and those determined by Orr et al. (2019), did not clearly resolve the phylogenetic position of the new species (Supplementary Figs 1 View Fig , 2 View Fig ), recovering a monophyletic relationship of the latter with a clade of “Elassodisca” and Prognatholiparis (bootstrap value: <50%) and “Melanura” (<50%) of Orr et al. (2019) in the ML and NJ trees, respectively. Therefore, the new species is here conservatively treated as a member of Careproctus , although future studies may not support this decision.
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.