
Ceraticelus Simon, 1884
|
publication ID |
https://doi.org/10.5281/zenodo.190335 |
|
DOI |
https://doi.org/10.5281/zenodo.6225639 |
|
persistent identifier |
https://treatment.plazi.org/id/997CC11F-FFC9-0101-FF78-1A4338ABE442 |
|
treatment provided by |
Plazi |
|
scientific name |
Ceraticelus Simon, 1884 |
| status |
|
Ceraticelus Simon, 1884 View in CoL View at ENA
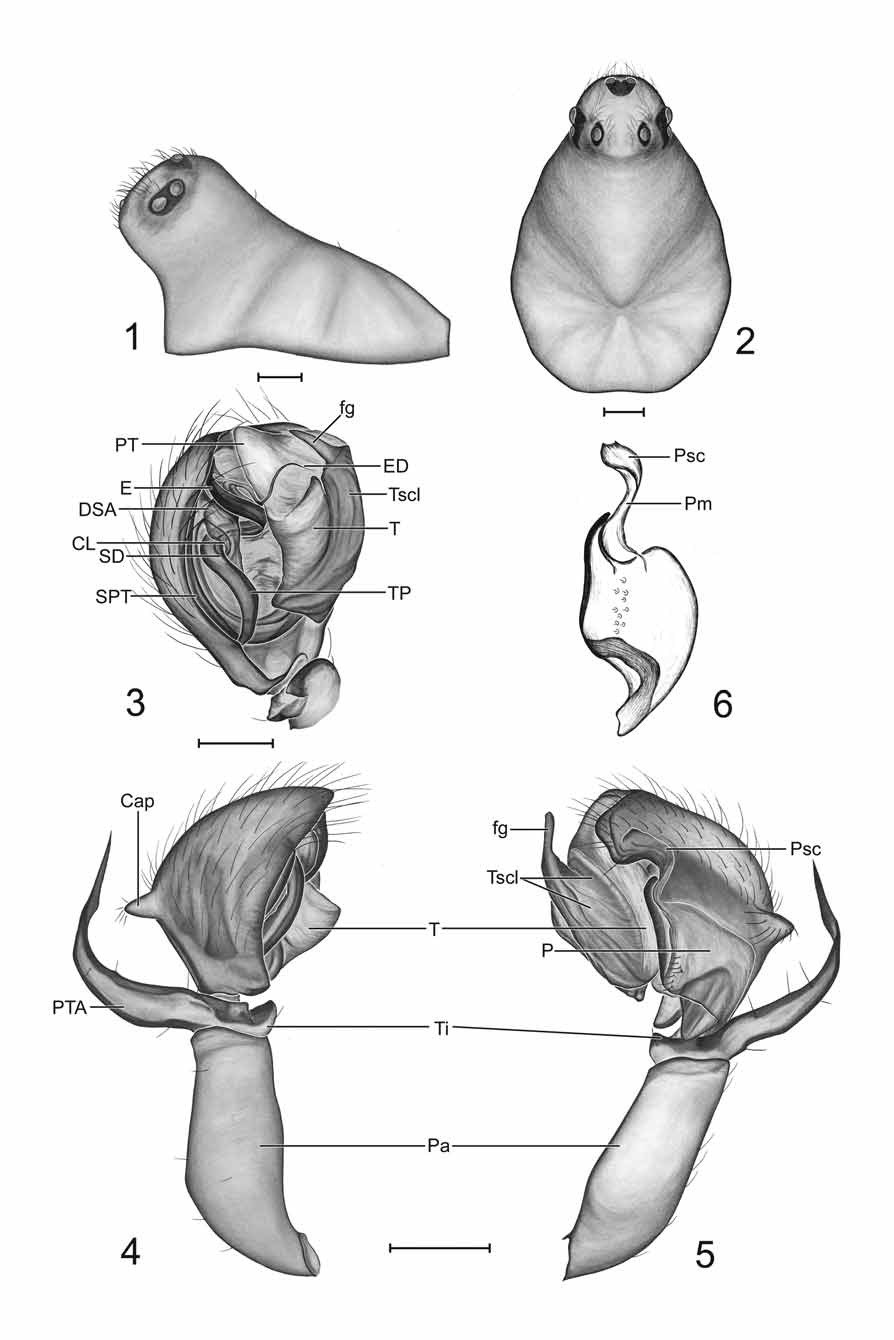
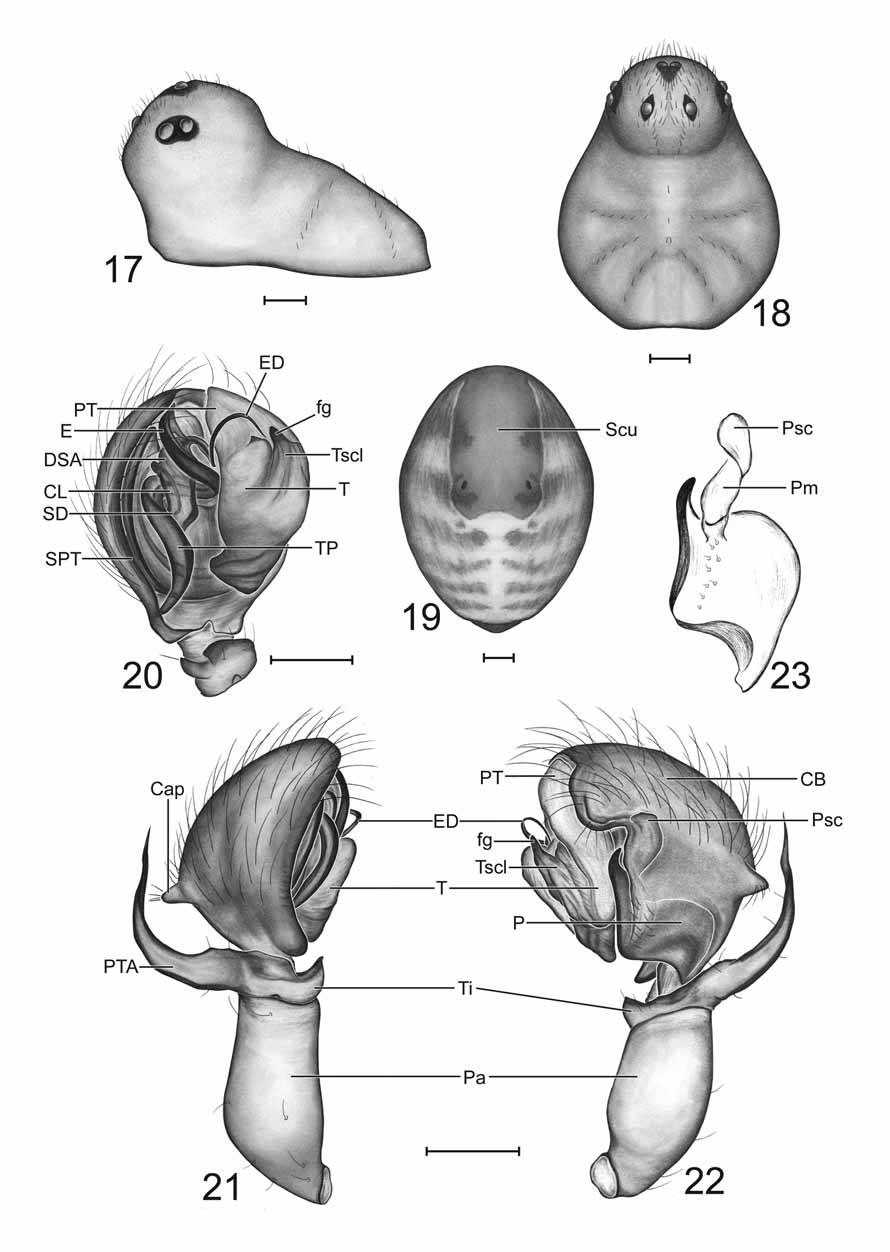
Type species: Erigone fissiceps O.P.- Cambridge, 1874 Simon (1884) proposed the genus Ceraticelus to house most of the species that Emerton (1882) had placed within Ceratinella Emerton. Simon did not consider these species to be congeneric with Theridium breve Wider , the species he then designated as the type of Ceratinella . He later ( Simon 1894) designated Ceraticelus fissiceps (O. P.-Cambridge) as the type of Ceraticelus . Essentially, the only character that then separated the two genera was the presence of a double curve of the fang of the chelicera in Ceratinella , “first concave and then convex without” ( Crosby & Bishop 1925).
Banks (1893) proposed the genus Idionella Banks for Ceratinella formosa Banks distinguishing it only “by the position of the horny shield”. Crosby (1905) placed Idionella in the synonymy of Ceraticelus stating that Banks’ character was “a character of less than specific value” given that in certain species the degree of development of this sclerite varies considerably between conspecifics of the same sex. Without explanation, Ivie (1967) removed Idionella from the synonymy of Ceraticelus and transferred C. anomalus Gertsch & Ivie , C. desertus Gertsch & Ivie , C. formosus (Banks) , C. guttatus Chamberlin & Ivie (= Idionella anamola (Gertsch & Ivie) , C. nesiotus Crosby , C. rugosus Crosby , C. titivillitium Crosby & Bishop , C. tuganus Chamberlin , and Grammonta sclerata Ivie & Barrows to the resurrected genus.
Millidge (1977) maintained that the double curvature of the cheliceral fang was probably of little significance given that Ceratinella scabrosa (Cambridge) in Europe have a single curve to the fang and that at least some of the North American species of Ceraticelus appeared to fall into Ceratinella although he did not specify to which species he was referring. Millidge further stated that in the European Ceratinella species the tibiae are armed with a single spine (also a single spine in the Nearctic species, C. brunnea Emerton ) and TmIV is present. Both Idionella species that we have examined, I. sclerata and I. nesiotus , possess tibial spines but lack TmIV and examined specimens of five Ceraticelus species, C. fissiceps (O. P.-Cambridge) (both sexes), C. emertoni (O.P.-Cambridge) (female), C. alticeps (Fox) (female), C. laetus (O. P.-Cambridge) (male; species very doubtfully belongs in Ceraticelus ), and C. phylax , lack tibial spines altogether as well as TmIV. In redescribing C. sibericus Eskov (= C bulbosus (Emerton)) from Poland, Kupryjanowicz (1994) stated that the species lacked tibial spines and TmIV as well as cheliceral stridulatory striae.
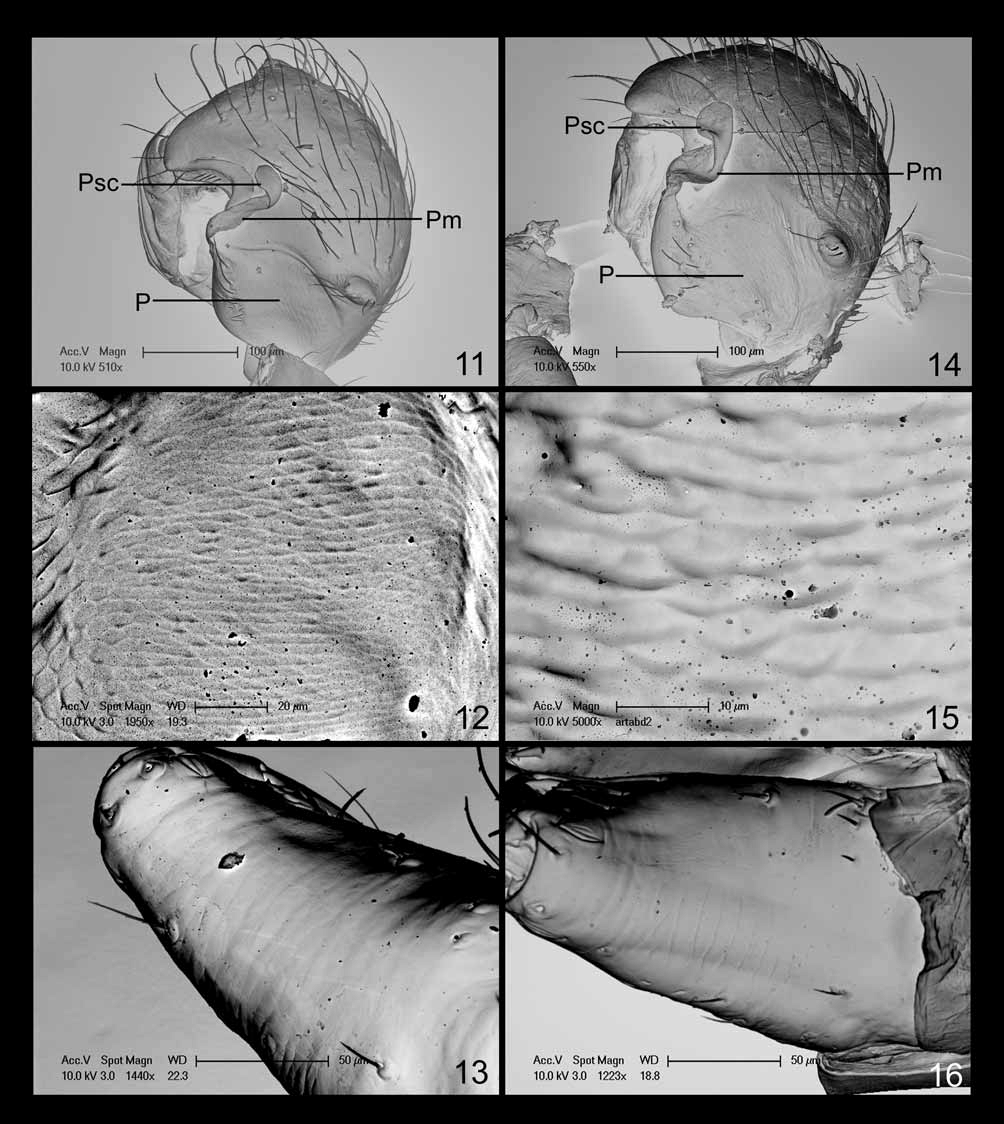
Millidge (1993) commented that the paracymbium “in some species of Ceraticelus Simon appears to have been produced by sclerotization of a laminar margin of the cymbium, with only a minimal degree of fission from the cymbium”; his illustration was of the cymbium of C. fissiceps , the type species. Upon examination of C. fissiceps specimens from AMNH (examined by either W. Ivie or C. R. Crosby), we found the paracymbium to be of the same form as in C. phylax and C. artemisiae sp. nov., a thin sclerite with incomplete fission from the cymbium (possibly connected by semi-membranous, weakly sclerotized tissue). In none of the species descriptions or redescriptions was there a mention of this type of paracymbium. In association with the paracymbia of C. fissiceps , C. artemisiae sp. nov., and C. phylax , there is also a very unique, previously overlooked structure in the form of a membranous bridge of tissue extending from the attachment point near the basal portion of the free distal finger-like projection of the paracymbium to the cymbium and terminating as a scale-like structure which lies on the surface of the cymbium, distad or opposite of the free hooked paracymbial tip (Pm, Psc, respectively: Figs 6 View FIGURES 1 – 6 , 11, 14 View FIGURES 11 – 16 , 23 View FIGURES 17 – 23 ). This two-part structure (membrane and scale) may well be a synapomorphy of Ceraticelus (the function of the structure and method of the scale attachment to the cymbium are presently unknown). In the expanded bulb (expansion during preservation), the curved apex of the paracymbium of C. artemisiae sp. nov. is very slightly rotated away from the edge of the cymbium (like the minute hand on a clock rotated away from the hour hand at 12:01AM) and clear tissue that appears somewhat sclerotized can be seen between the cymbial and paracymbial margins. This slight separation of paracymbium and cymbium indicates that there is at least some degree of fission between the structures. The flattened paracymbial scale (Psc: Figs 11, 14 View FIGURES 11 – 16 ) becomes detached from the cymbium in the expanded bulb of C. artemisiae (presumably also in C. phylax and C. fissiceps ).
Further examination of the bulb of C. fissiceps (at 200X) revealed a sclerotized column and what was presumed to be a rather short sclerotized distal suprategular apophysis which appeared to support the embolus somewhere near the break point of the ejaculatory duct from the embolic ribbon. The tip of the ejaculatory duct, as in C. phylax , was found to rest in the cradle formed between the tegulum and protegulum. We detected neither an embolic membrane nor the white membranous lobe (conductor) that Crosby & Bishop (1925) mentioned as lying just inside the tip of the (bezel) protegulum (a more detailed examination involving dissection of the male bulb was not a provided option for the borrowed material).
The unique form of the paracymbium, with the associated paracymbial membrane and scale, and strong similarity in bulb conformation, chaetotaxy, and arrangement of abdominal sclerites shared by C. fissiceps , C. phylax , and C. artemisiae sp. nov., indicate that the species are congeneric. The presence in C. fissiceps of cuticular plectra on coxae IV, the apparent lack of functional cheliceral striae (viewed at 200X), and an elongated palpal patella further support the taxonomic placement of C. phylax and C. artemisiae sp. nov.
We postulate that C. phylax and C. artemisiae sp. nov. are probable sibling species based on the nearly identical conformation of the male bulbs and respective morphologies of the tibial apophysis, cymbium, and paracymbium, the states of the latter three characters differing from all other congeneric males, and on the identical convolution of the female copulatory ducts.
Diagnosis. The incomplete fission of the cymbium and paracymbium, with the associated paracymbial membrane and scale (which may be synapomorphic for the genus) separates Ceraticelus from those genera with which it has been confused, Ceratinella and Idionella , which have intersegmental, ‘J’ shaped paracymbia ( Hormiga 2000) without associated structures. Characters in combination which then provisionally define the genus are the incomplete fission of the cymbium and paracymbium (P: Figs 11, 14 View FIGURES 11 – 16 ), with the associated paracymbial membrane and scale (Pm, Psc, respectively: Figs 11, 14 View FIGURES 11 – 16 ), male bulb with a spiraling type of embolus with the ejaculatory duct breaking away from the embolic ribbon preapically and spiraling ventrally, long tailpiece, sclerotized distal suprategular apophysis which apparently supports basal portion of the ejaculatory duct, sclerotized column, similarity in arrangement of abdominal sclerites (especially in male), an elongated palpal patella and short palpal tibia (usually with a long tibial apophysis) in males, and the lack of tibial spines (or extreme reduction of) and TmIV in both sexes and cephalic sulci in males.
| AMNH |
American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |