
Mecynogea infelix ( Soares & Camargo, 1948 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4415.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:5E60ED09-4D71-4F64-8165-3AD59B30FE3A |
|
DOI |
https://doi.org/10.5281/zenodo.5995857 |
|
persistent identifier |
https://treatment.plazi.org/id/9A05197E-FF83-FFD7-FF78-78FF66460FCB |
|
treatment provided by |
Plazi |
|
scientific name |
Mecynogea infelix ( Soares & Camargo, 1948 ) |
| status |
|
Mecynogea infelix ( Soares & Camargo, 1948) View in CoL revalidated
Figs 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8
Wixia infelix Soares & Camargo, 1948: 378 , fig. 35 (female holotype from left side of Rio das Mortes, Xavantina, Mato Grosso, Brazil, [18°14'58"S, 43°36'1"W], 6.II.1947, H. Sick leg., deposited in MZSP 1300, examined). Brignoli 1983: 281. Mecynogea infelix View in CoL — Levi 1993: 56
Mecynogea bigibba Simon, 1903 View in CoL — Levi 1997: 222 –224 (Syn).
Mecynogea chavona Levi, 1997: 230 View in CoL , figs 65–66, map 2D ( Male holotype from Finca Chenevo, 20 km N Río Muco, 20 km S El Porvenir, ca. 170 m, Depto. Meta, Colombia, [ 4°25'18.97"S, 72°0'53.93"W], 1979, W. Eberhard leg., deposited in MCZ 20767, examined by photograph). New synonymy. GoogleMaps
Mecynogea bigibba Simon, 1903 View in CoL — Levi 1997: 222 –224, figs 10–14 (in part, some females).
Mecynogea lemniscata (Walckenaer, 1849) View in CoL — Levi 1997: 227 figs 46–48 (misidentified in part, only some females).
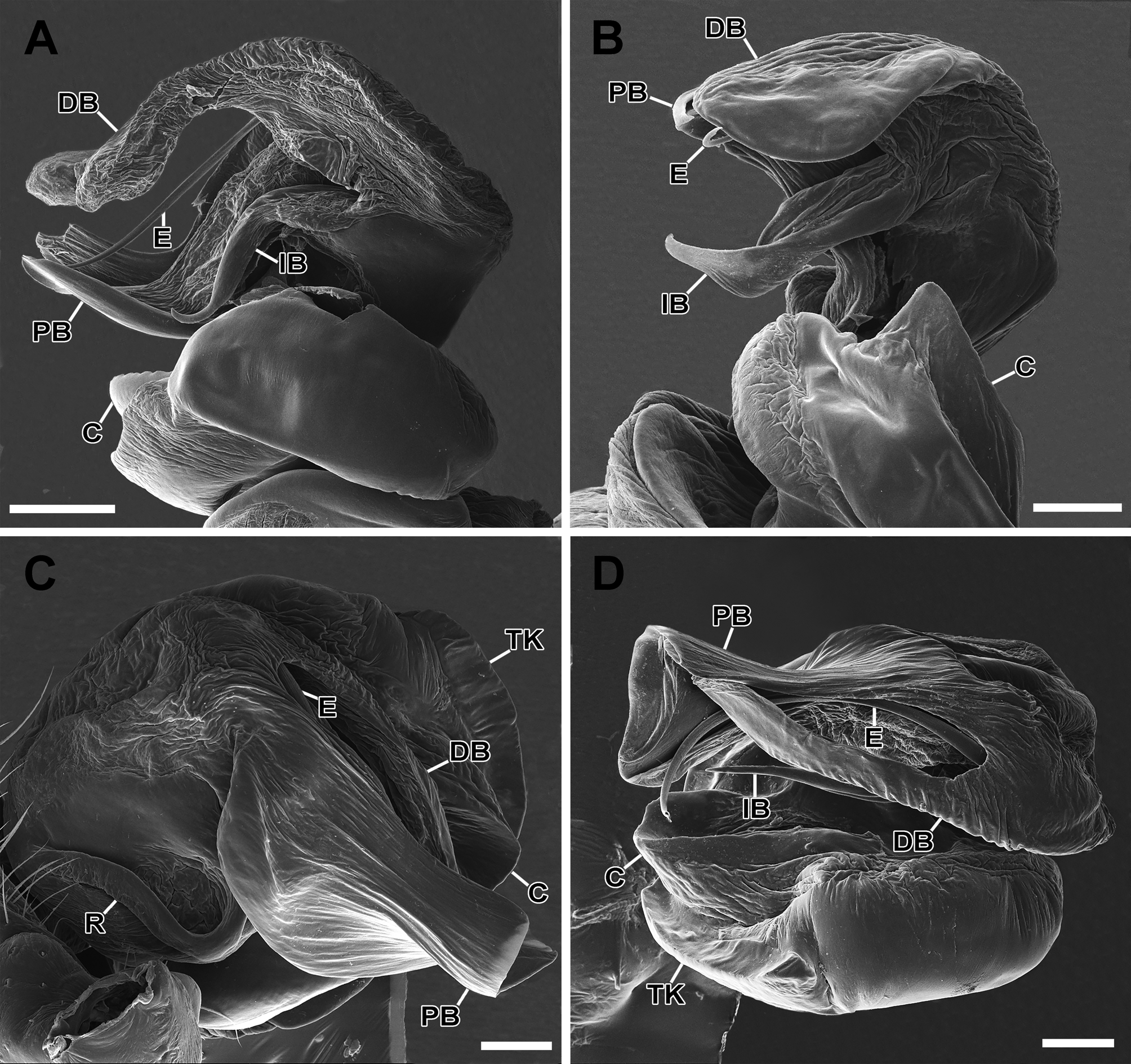
Diagnosis. Males and females of M. infelix can be distinguished from other species of the genus by the lack of a median black line and lateral black bands on the carapace ( Fig. 1A, B View FIGURE 1 ). Males of M. infelx resemble the males of M. lemniscata and M. sucre Levi 1997 , by the proximal branch of the terminal apophysis covering the tip of the distal branch of the terminal apophysis in mesal view ( Fig. 1C View FIGURE 1 , Levi, 1997: fig. 38). It can be separated from the males of M. lemniscata and M. sucre by the rounded tip of the proximal branch of the terminal apophysis ( Figs 1C View FIGURE 1 , 2A View FIGURE 2 ), which is truncated in M. lemniscata ( Fig. 14C View FIGURE 14 ), and acuminate in M. sucre ( Levi, 1997: fig. 38). M. infelix also can be differentiated by the less developed tegular keel ( Figs 1C View FIGURE 1 , 2A View FIGURE 2 , 3A View FIGURE 3 ), which is well developed in M. lemniscata ( Fig. 14C, D View FIGURE 14 ). It can also be diagnosed by the wide proximal branch of the terminal apophysis ( Levi, 1997). Females are most similar to M. lemniscata based on the shape of the epigynum, which have a deep cavity created by the epigynum hood in both species. M. infelix can be distinguished by the presence of a strong, sclerotized median septum on the posterior cavity of the epigynum ( Fig. 7C View FIGURE 7 ; see also Levi 1997: figs 11, 47), which is membranous in M. lemniscata . It can also be distinguished from M. leminiscata and other species of the genus by the presence of a fracture line on the epigynum hood ( Figs 4A–C View FIGURE 4 , 5A, B View FIGURE 5 , 6B, D, E View FIGURE 6 , 7A–D View FIGURE 7 ).
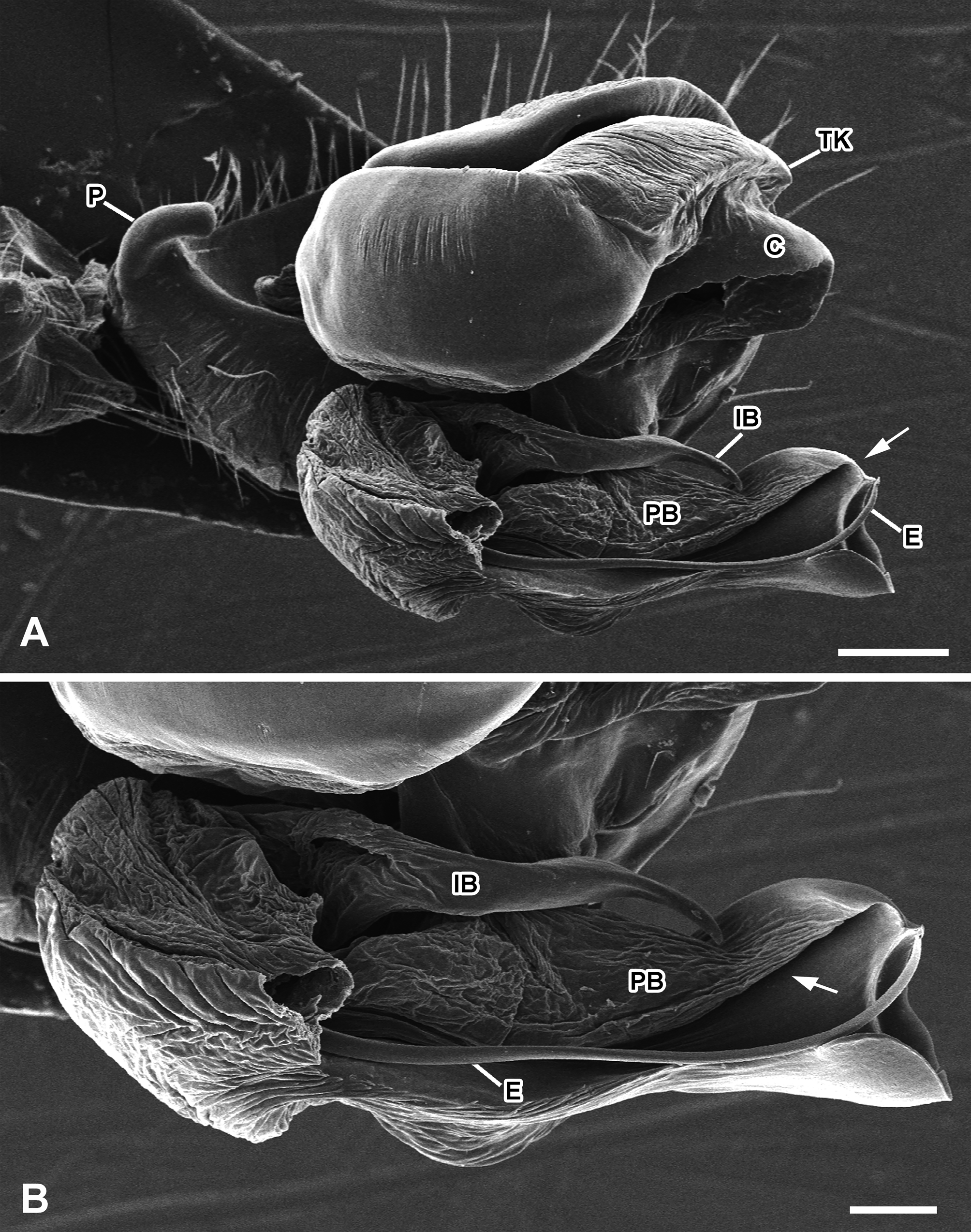
Description. Male. Colour and measurements described by Levi (1997: 230), under Mecynogea chavona . Male palp (based on specimen from Santana do Riacho, Minas Gerais, Brazil, UFMG 1739): terminal apophysis with a very developed and sclerotised proximal branch, which covers the apex of the distal branch in mesal view ( Figs 1C View FIGURE 1 , 2A View FIGURE 2 ). Proximal branch with a longitudinal depression holding the embolus, which is long and very thin ( Figs 2C View FIGURE 2 , 3A–B View FIGURE 3 ). Conductor well developed, strongly sclerotized and fused to ventral margin of tegulum ( Figs 1D View FIGURE 1 , 2A–B View FIGURE 2 , 3A View FIGURE 3 ). Internal branch of terminal apophysis elongated, gently sinuous and acuminated apically, hidden by the terminal apophysis branches ( Fig. 3A, B View FIGURE 3 ). Radix translucent and very poorly sclerotized ( Figs 1C View FIGURE 1 , 2A, B View FIGURE 2 ). Paracymbium small, apically rounded ( Fig. 3A View FIGURE 3 ).
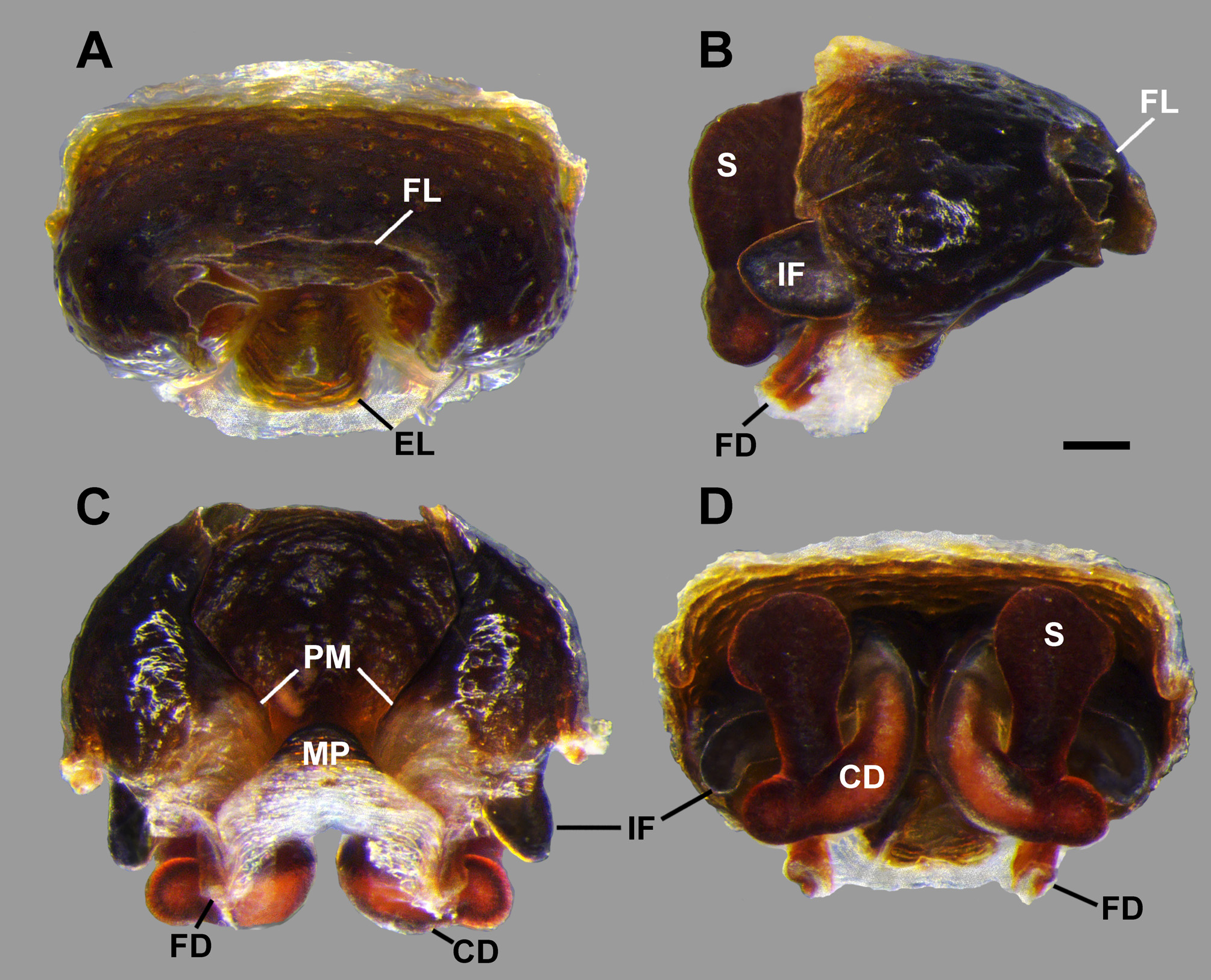
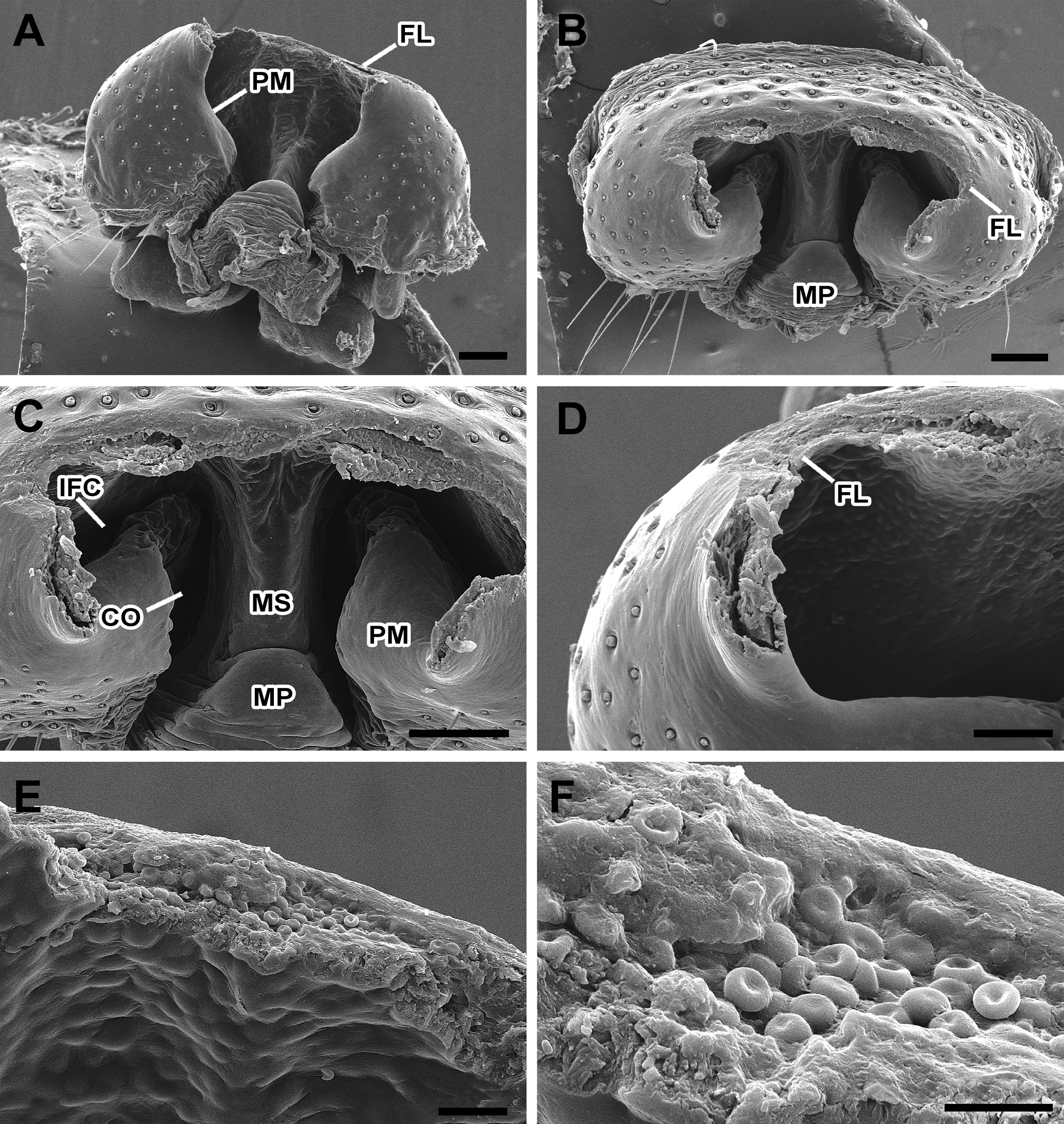
Female from Serra da Calçada, Minas Gerais, Brazil (IBSP 63237). Carapace orange without black bands ( Fig. 1A View FIGURE 1 ). Chelicerae proximally orange, gradually darker distally. Endites and labium dark brown proximally and white distally. Sternum orange with dark lateral blots. Palp light orange. Legs orange, first and second femur with a dorsal, longitudinal black line ( Fig 1A View FIGURE 1 ). All femora orange, darker ventrally. Coxae orange, with black markings on the second, third and fourth. Opisthosoma with a pair of anterior humps and an elaborated colour pattern, which shows a pair of black anterior markings; a longitudinal, median black mark with a transverse black bar, and two posterior black blots in a longitudinal red band with a trident shape pattern ( Fig. 1A View FIGURE 1 ). The dorsolateral and lateral sides show a reticulated black background with white sinuous stripes. Venter bearing black stripes, with a pair of longitudinal white bands and four white markings around the spinnerets. Total length 9.60; carapace 3.80 long, 2.90 wide in the thoracic area, 1.30 wide after the posterior median eyes. Femur I length 5.0, patella 1.50, tibia 3.70, metatarsus 4.70, tarsus 1.50. Patella II 1.50, III 1.00, IV 1.40. Tibia II 3.10, III 1.5, IV 2.90. Epigynum projected ventrally, atrium with large, slit-shaped posterior openings. The posterior margin of the hood is folded, entering the atrium, forming an inner fold ( Figs 4B, D–E View FIGURE 4 , 5B–D View FIGURE 5 , 8A–C View FIGURE 8 ) creating the cavity of the inner fold visible in the ventral view ( Fig. 7C View FIGURE 7 ). Epigynum shows a sclerotized median septum that originates on median plate. Copulatory openings situated at the sides of median septum ( Fig. 7C View FIGURE 7 ). Spermathecae divided in two receptacles ( Fig. 8A, B View FIGURE 8 ), one small proximal and another larger and sock-shaped ( Fig. 8C View FIGURE 8 ). Copulatory ducts emerge medially between larger spermathecae receptacles, folded, passing dorsally to the large receptacles ( Fig. 8A, B View FIGURE 8 ). Both spermathecae receptacles show several small and large pores ( Fig. 8D View FIGURE 8 ). Fertilization ducts connected proximally to spermathecae, near proximal receptacle ( Fig. 8C View FIGURE 8 ).
Variation. Three of the seven females from Serra do Cipó have an entire epigynum with a rounded hood, which hides the atrium with the copulatory openings. The remaining females, from Brasilândia de Minas, Serra da Moeda and four specimens from Serra do Cipó lack the apex of the epigynum. The total length of females varies from 7.20 to 10.13 (n, 4).
Remarks. M. infelix was described based on a single female from Mato Grosso, central-western Brazil ( Soares & Camargo 1948), and was considered by Levi (1997) as a junior synonym of M. bigibba based on the examination of the both species’ type material. In this same study, he described M. chavona based on a single male from Meta, Colombia.
We have found on the UFMG collection a sample of seven males and seven females collected together (UFMG 1739). The males match perfectly the diagnosis of M. chavona . However, the females show a very weird genital pattern. Three of them have an epigynum similar to those of M. lemniscata ( Levi, 1997: figs 46–48) and of the holotype of M. infelix ( Levi, 1997: figs 13–14), which have a deep cavity created by the epigynum hood. However, they differ from M. lemniscata by having a sclerotized medium septum on the epigynum, and by lacking the median black line and the black bands on the carapace edges. The holotype of M. infelix shares the same features with those three specimens. The remaining four females apparently have their epigynum damaged, missing the apex, making them very similar to some females illustrated by Levi (1997: figs 10–12) as M. bigibba . Those specimens considered by Levi as a variation of M. bigibba matched better the holotype of M. infelix than the holotype of M. bigibba by having a posterior atrium in the epigynum rather than the small posterior bulge seen in M. bigibba , leading us to reconsider the synonymy between M. bigibba and M. infelix .
Additionally, because we have found seven males that match the description made by Levi (1997), which are identical to the holotype of M. chavona , and were collected together with seven females that match the M. infelix holotype, we concluded that M. chavona should be considered a junior synonym of M. infelix .
Natural History. Several female specimens examined for this study have mutilated epigyna, in which the apex has apparently been removed ( Figs 5A–C View FIGURE 5 , 7A–D View FIGURE 7 ). The break in the epigynum coincides with an apparent fracture line ( Fig. 4A–C View FIGURE 4 ). The mutilation of female epigynum during or after copulation has been reported for other araneid genera, and has been seen as a male strategy to prevent subsequent female mating events with other males ( Mouginot et al. 2015, Nakata 2016). We think this explanation could possibly apply to M. infelix . We also noted cell clumps in the epigynum surface of different specimens, particularly on the fracture line ( Figs 6C, F–H View FIGURE 6 , 7E–F View FIGURE 7 ). These cells resisted the enzymatic cleaning treatment, indicating a chitin cell wall. The cells in our images are similar to the conidia of Beauveria bassiana , an entomopathogenic fungus ( Wakefield 2006: fig 3a–d). It is unclear at this point whether these apparent fungal cells have any importance for the female epigynum mutilation, or if they simply spread on the fracture line.
Regarding the male copulatory apparatus, the shape and position of the proximal branch of the terminal apophysis suggest it could act as a functional conductor during copulation. Additionally, the remarkable size and sclerotization, as well as the folded shape of this sclerite ( Fig. 2C View FIGURE 2 ) could indicate it is somehow related to female epigynum mutilation.
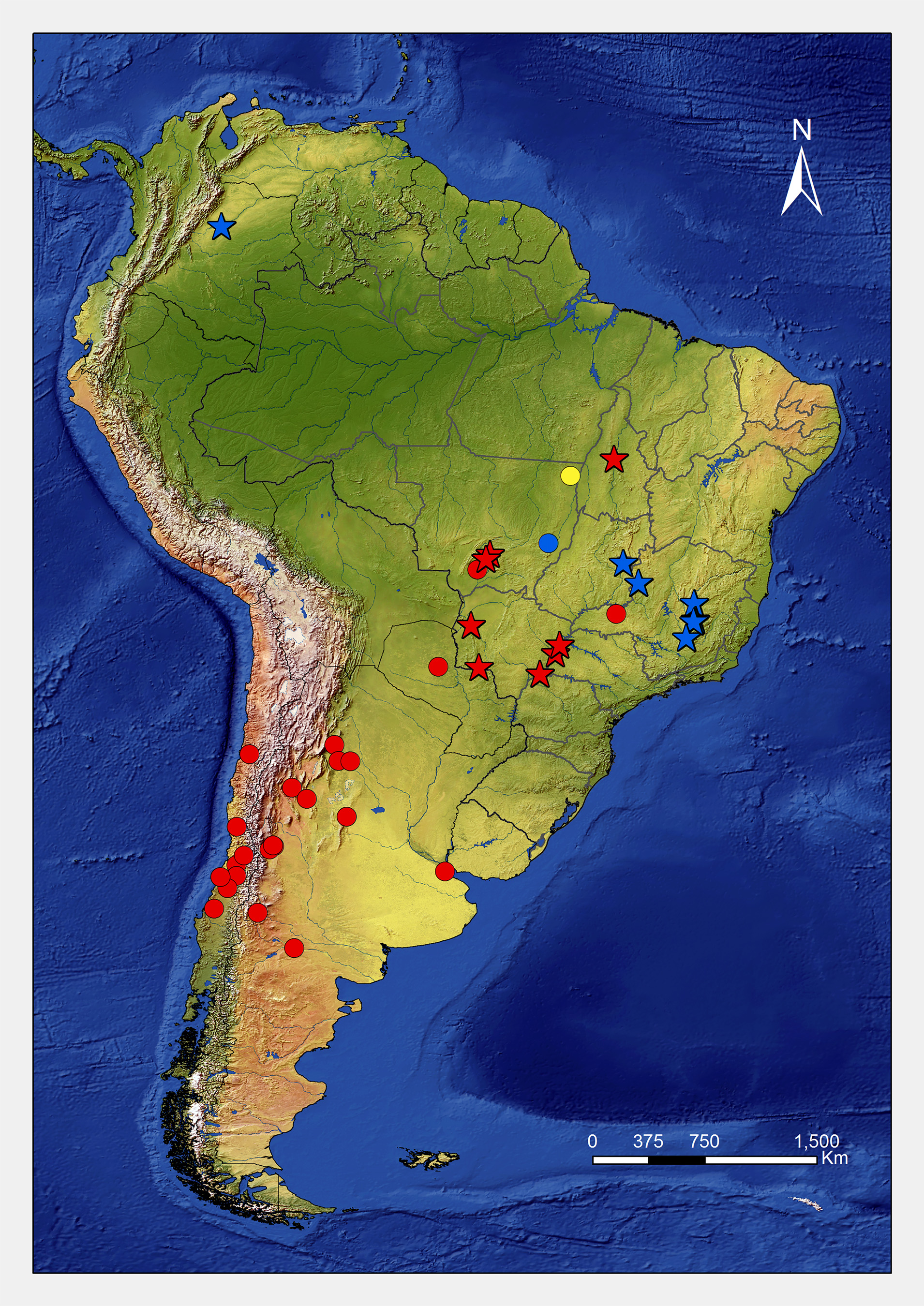
Distribution. This species is distributed mostly in the Neotropical savannas, including the Colombian llanos, which represent the northernmost record of the species. In Brazil, it can be found in the Distrito Federal, Minas Gerais, and Mato Grosso ( Fig. 16 View FIGURE 16 ).
Additional material examined. BRAZIL: Distrito Federal: Brasília, São Francisco, Lago Paranoá [ 15°50'41.16"S, 47°50'17.75"W], 1038 m, I.1978, 1 ♀, ( MZSP 70592 View Materials ) GoogleMaps ; Minas Gerais: Brasilândia de Minas, Fazenda Brejão [ 17°1'9.72"S, 46°54'59.81"W], 4.IV.2004, R.A.F. Redondo & F.A. Perini leg. GoogleMaps , 5♂ 1♀ ( UFMG 1536 View Materials ); Diamantina [ 18°14'58"S, 43°36'1"W], 1236 m GoogleMaps , 2♀ 1 juv. ( MNRJ 2331 View Materials ); Moeda, Serra da Moeda [ 20°20'1.16"S, 44°3'18.82"W], 809 m GoogleMaps , 1♀, 20.V.2006, E.S.S. Álvares leg. (IBSP 63237); Santana do Riacho, Parque Nacional da Serra do Cipó [ 19°20'57"S, 43°37'10.16"W], 822 m, I.2004, E.S.S. Álvares leg., 5♂ 2♀ ( UFMG 1739 View Materials ) GoogleMaps ; 1♀ (UFMG 19542); 1♀ (UFMG 19543); 1♂ (UFMG 19890); 1♀ (UFMG 19891); 1♀, (UFMG 19892); 1♂ ( UFMG 19893 View Materials ), ditto, Portaria Palácio ( 19°15'0"S, 43°31'0"W), 1200 m, 10.XII.2001, E.S.S. Álvares & E.O. Machado leg. GoogleMaps , 1♂ (UFMG 1364).





| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mecynogea infelix ( Soares & Camargo, 1948 )
| Martins, Pedro H. & Santos, Adalberto J. 2018 |
Wixia infelix
| Soares & Camargo, 1948 : 378 |
| Brignoli 1983 : 281 |
| Levi 1993 : 56 |
Mecynogea bigibba
| Levi 1997 : 222 |
Mecynogea chavona
| Levi, 1997 : 230 |
Mecynogea bigibba
| Levi 1997 : 222 |
Mecynogea lemniscata (Walckenaer, 1849)
| Levi 1997 : 227 |