
Limnadopsis tatei Spencer & Hall, 1896
|
publication ID |
https://doi.org/ 10.5281/zenodo.210812 |
|
DOI |
https://doi.org/10.5281/zenodo.5619656 |
|
persistent identifier |
https://treatment.plazi.org/id/9A5D7F26-FFD7-FFDE-7294-FDB5FD29F860 |
|
treatment provided by |
Plazi |
|
scientific name |
Limnadopsis tatei Spencer & Hall, 1896 |
| status |
|
Limnadopsis tatei Spencer & Hall, 1896 View in CoL
( Figs. 1 View FIGURE 1 c, d, 4)
Limnadiopsis tatei: Daday 1925: 181 –183
Limnadiopsisum tatei : Novojilov 1958: 104, Brtek 1997: 58 Limnadopsis cf. tatei View in CoL ‘Titanic’: Schwentner et al. 2011
Material examined: New South Wales: 23, 2Ƥ (AM P. 85168, P. 85169, AM P.86375, P.86376, Genbank JN698540 View Materials , JN698541 View Materials , JN698550 View Materials , JN698551 View Materials ), Beverley's Pool, Bloodwood Station, 29°32'12.0''S, 144°51'16.1''E, 19.ii.2010; 13, 1Ƥ (AM P.85162, AM P.85163, GenBank JN698534 View Materials , JN698535 View Materials ), Gidgee Lake, Bloodwood Station, 29°33'10.4''S, 144°50'12.7''E, 19.ii.2010; 43, 1Ƥ (AM P.85164, AM P.85165, P.86372 -P.86374, GenBank JN698536 View Materials , JN698537 View Materials , JN698547 View Materials - JN698549 View Materials ), homestead swamp 1, Bloodwood Station, 29°31'31.5''S, 144°53'27.7''E, 19.ii.2010; 13 (AM P.85170, GenBank JN698542 View Materials ), Upper Crescent Pool, Bloodwood Station, 29°32'33.6''S, 144°52'16.5''E, 19.i.2010; 23, 1Ƥ (AM P.85166, AM P.85167, AM P.86377, GenBank JN698538 View Materials , JN698539 View Materials , JN698552 View Materials ), Roskos Paleolake, Bloodwood Station, 29°27'42.9''S, 144°48'12.5''E, 19.ii.2010; 1Ƥ (AM P.86315, GenBank JN698504 View Materials ), Lismore Bore, Muella Station, 29°31'50.7''S, 144°59'28.1''E, 19.i.2010; 1Ƥ (AM P.85171, GenBank JN6985439), Lower Lake Eliza, Muella Station, 29°25'28.9''S, 145°03'41.8''E, 20.ii.2010; 13 (AM P.86385, GenBank JN698560 View Materials ), Quandong Swamp, Tredega Station, 29°27'45.4''S, 144°51'06.6''E, 23.ii.2011; 13 (AM P.85161, GenBank JN698257 View Materials ), pool east of Lake Lauradale, 29°51' 22''S, 145°38' 49''E, 23.ii.2009; South Australia: 53, 5Ƥ (AM P.85864 – AM P. 85873; GenBank JF966719 View Materials - JF966728 View Materials ), vegetated stony dugout 34 km north of Marla, 27°05'26.8''S, 133°28'16.2''E, 10.iii.2011; Queensland: 33, 2Ƥ (AM P.86378 - AM P.86382; Gen- Bank JN698553 View Materials - JN698557 View Materials ), beefwood grassy swamp, 28°50'34.5''S, 143°53'47.3''E, 26.ii.2011; Western Australia: 13 (AM P.87801; GenBank JQ424942 View Materials ), Lake Carey, 08.iii.2011 (collected by Fiona Taukalis).
Short description and variability. The morphological variability of L. tatei will only be described on the basis of specimens for which COI data are available. The most important features are summarized in Table 2 View TABLE 2 . Timms (2009) has already described the morphological variability of L. tatei , but specimens of L. paratatei may have been included in that description.
Carapace varies in its overall shape ( Fig. 1 View FIGURE 1 c, d). Dorsal margin is straight or slightly convex in both genders. Serration at the dorsal margin is usually well developed with strong, long ‘spines’ extending from most growth lines. In some specimens the serration can be reduced to small ‘spines’ or completely absent, probably due to abrasion. Growth lines are always well expressed and regularly arranged with distance between growth lines increasing towards the ventral margin. The outermost growth lines (usually two or three) are crowded at the carapace’s margin in some specimens.
First antenna is made up of 7–13 lobes ( Fig. 4 View FIGURE 4 a, b) and can be so short that it does not reach the base of the second antenna, or it can extend to the third antennomere.
Second antenna has 11–15 antennomeres, and in every specimen this number was greater than that of the lobes of the first antenna.
Telson has 9–14 spines (Spencer & Hall reported up to 15) which are always irregularly sized and spaced ( Fig. 4 View FIGURE 4 e, f). In most specimens a perceptibly larger gap separates the first spines from the last three to five spines. The last spine is greatly enlarged and bears one to four of the preceding spines at its base. In all males the last enlarged spine on the right side of the telson is bent at approximately 90° whereas the spine on the left side is fairly straight ( Fig. 4 View FIGURE 4 e). In females the difference is less pronounced, with the spine on the right side being straighter than in males ( Fig. 4 View FIGURE 4 f). This difference between males and females can also be seen in the drawings by Spencer & Hall (1896). Differences between the two sides of the telson are also present in other species of Limnadopsis (e.g. L. parvispinus , L. minuta Timms 2009 , L. occidentalis and L. paradoxa as seen in Timms 2009 Fig. 2 View FIGURE 2 ) and are not specific to L. tatei .
L. tatei Spencer & L. tatei (male) L. tatei (female) L. paratatei sp. nov. L. paratatei sp. nov
Hall 1896 (male) (female) growth lines 10–13; 10* 9–15 (11–12) 8–12 (10) 11–18 (15) 10–16 (14) Furcal rami bear 6–18 setae which cover the first third to two-thirds of each ramus ( Fig. 4 View FIGURE 4 e, f). These are followed by one to four spines, the last of which is always enlarged (where only one spine is present, it is enlarged). The larger spine is situated at about two-thirds or three-quarters of the length of the furcal ramus. The last third or quarter of the furcal ramus is covered in numerous denticles. This part of the ramus is always strongly bent, while the rest is either straight or only weakly bent. The tip of the movable finger of the male clasper features a single rounded scale without a ventral ledge ( Schwentner et al 2011).
Genetic diversity of COI within L. tatei is low: we found only 0.0–0.8 % uncorrected p-distance and K2P corrected distance between specimens from east and central Australia and about 1.5 % distance to one Western Australian specimen. No genetic structure was evident within the species’ distribution range in central and eastern Australia (Schwentner et al. in press).
Distribution. L. tatei occurs in large parts of New South Wales and southern Queensland (Murray-Darling Basin and catchment of the Bulloo River) as well as in northern South Australia (Lake Eyre Basin) and Western Australia. Timms (2009) further reported L. tatei in south-eastern South Australia and Weeks et al. (2009) in the Northern Territory. L. tatei thus seems to inhabit large parts of arid and semi-arid Australia. However, these other distribution records may in some instances have mistaken L. tatei for L. paratatei sp. nov. or even other L. tatei - like species. The same is true for other sites within the catchment of the Paroo River reported by Timms (2009) and Timms & Richter (2002).
Remarks. Spencer & Hall first described L. tatei in 1896 from a group of eight specimens collected between Charlotte Waters and Oodnadatta (northern South Australia). As they did not designate a holotype, Timms (2009) designated a neotype (as the specimen is believed to stem from the original type series it should properly be termed a lectotype) and extended the original description of L. tatei . Specimens identified as L. tatei could consistently be delineated into two species (provisionally termed L. cf. tatei ‘Titanic’ and L. cf. tatei ‘Carter’s’) in an integrative taxonomic approach by Schwentner et al. (2011). A comparison of those specimens with the original description of L. tatei and the additional data provided by Timms (2009) clearly identified L. cf. tatei ‘Titanic’ as being L. tatei Spencer & Hall, 1896 , as suggested by Schwentner et al. (2011). Characteristic differences between L. tatei and the new species L. paratatei will be discussed below.

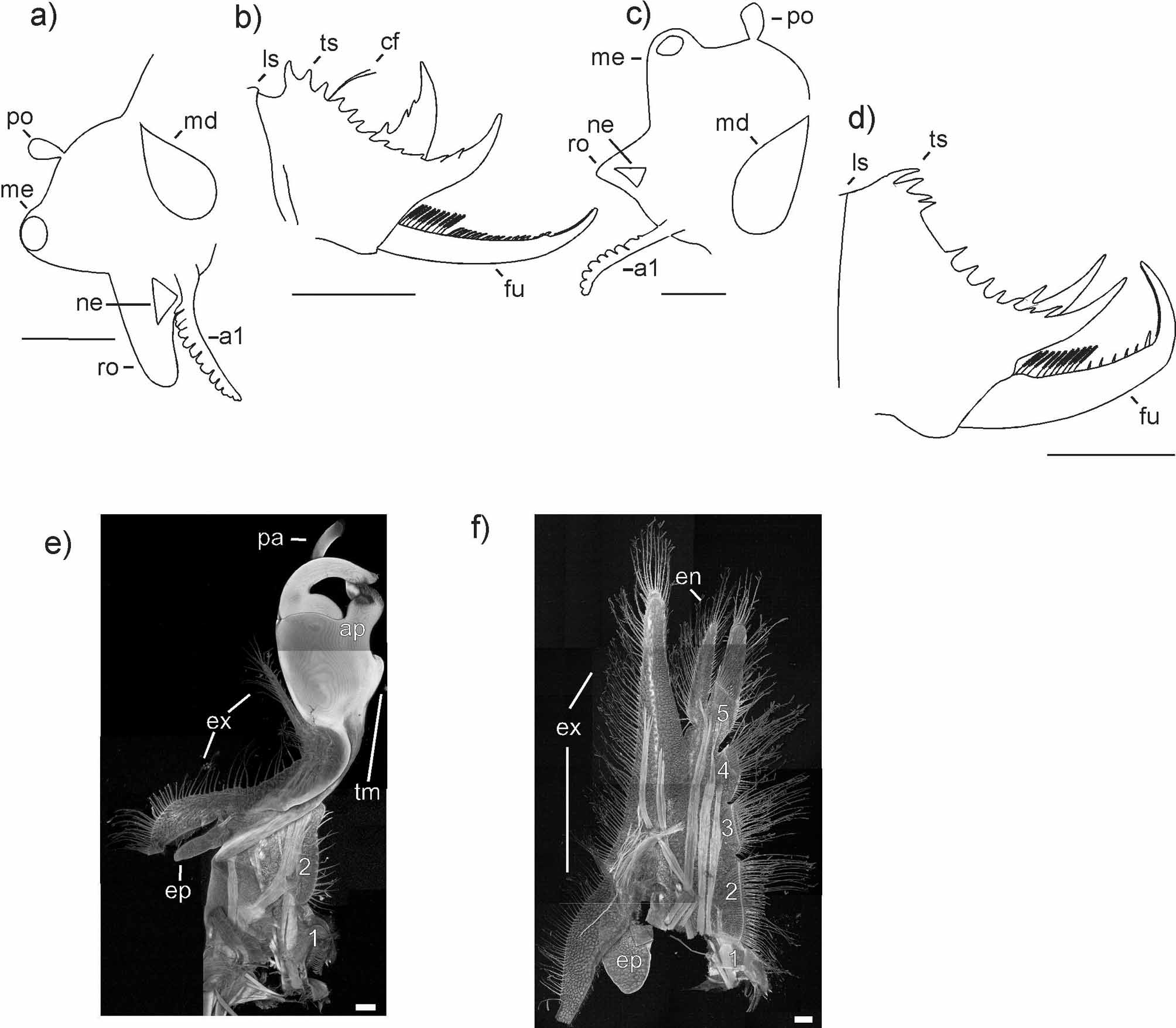

TABLE 2. Comparison of morphological characteristics of the species Limnadopsis tatei and L. paratatei sp. nov. For L. tatei, details of the original description by Spencer & Hall (1896) are also given. Ranges and median values (in brackets) are given for males and females of both species. * based on the description of the male lectotype chosen and described by Timms (2009); ** based on Schwentner et al. (2011)
| dorsal serration on car- strong apace | strong in most speci-strong in most spec- weaker, short mens imens | weaker, short |
|---|---|---|
| trunk segments 26; 26* | 26–27 (27) 26–27 (27) 24–25 (25) | 24–25 (25) |
| 1st antenna 10* | 7–13 (11) 7–11 (9) 6–12 (8–9) | 5–8 (7) |
| 2nd antenna 14; 15* | 12–15 (14) 11–15 (14) 11–17 (15) | 12–17 (14) |
| spines on telson 10 –15; 11* | 9–14 (12) 9–14 (11) 14–21 (18) | 15–21 (19) |
| setae on furcal ramus 10* | 6–18 (12–13) 8–12 (10) 5–14 (8) | 6–12 (8) |
| spines on furcal ramus 1* | 1–4 (1) 1–3 (2) 1–4 (1–2) | 1–5 (2) |
| eggs – | – irregular ridges – | polygonal ridges sur- rounding oval ridge |
| tip of movable finger – of male clasper | round without ledge, – round with ledge, length 20–44 µm ** length 40–53µm ** | – |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Limnadopsis tatei Spencer & Hall, 1896
| Schwentner, Martin, Timms, Brian V. & Richter, Stefan 2012 |
tatei
| Brtek 1997: 58 |
| Novojilov 1958: 104 |
Limnadiopsis tatei:
| Daday 1925: 181 |