
Sulaplax, Naruse, Tohru, Ng, Peter K. L. & Guinot, Danièle, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181452 |
|
DOI |
https://doi.org/10.5281/zenodo.6233822 |
|
persistent identifier |
https://treatment.plazi.org/id/9C7287B0-9944-FFAA-C195-D071FE7EFEC1 |
|
treatment provided by |
Plazi |
|
scientific name |
Sulaplax |
| status |
gen. nov. |
Sulaplax View in CoL gen. nov.
Type species. Sulaplax ensifer spec. nov. by present designation.
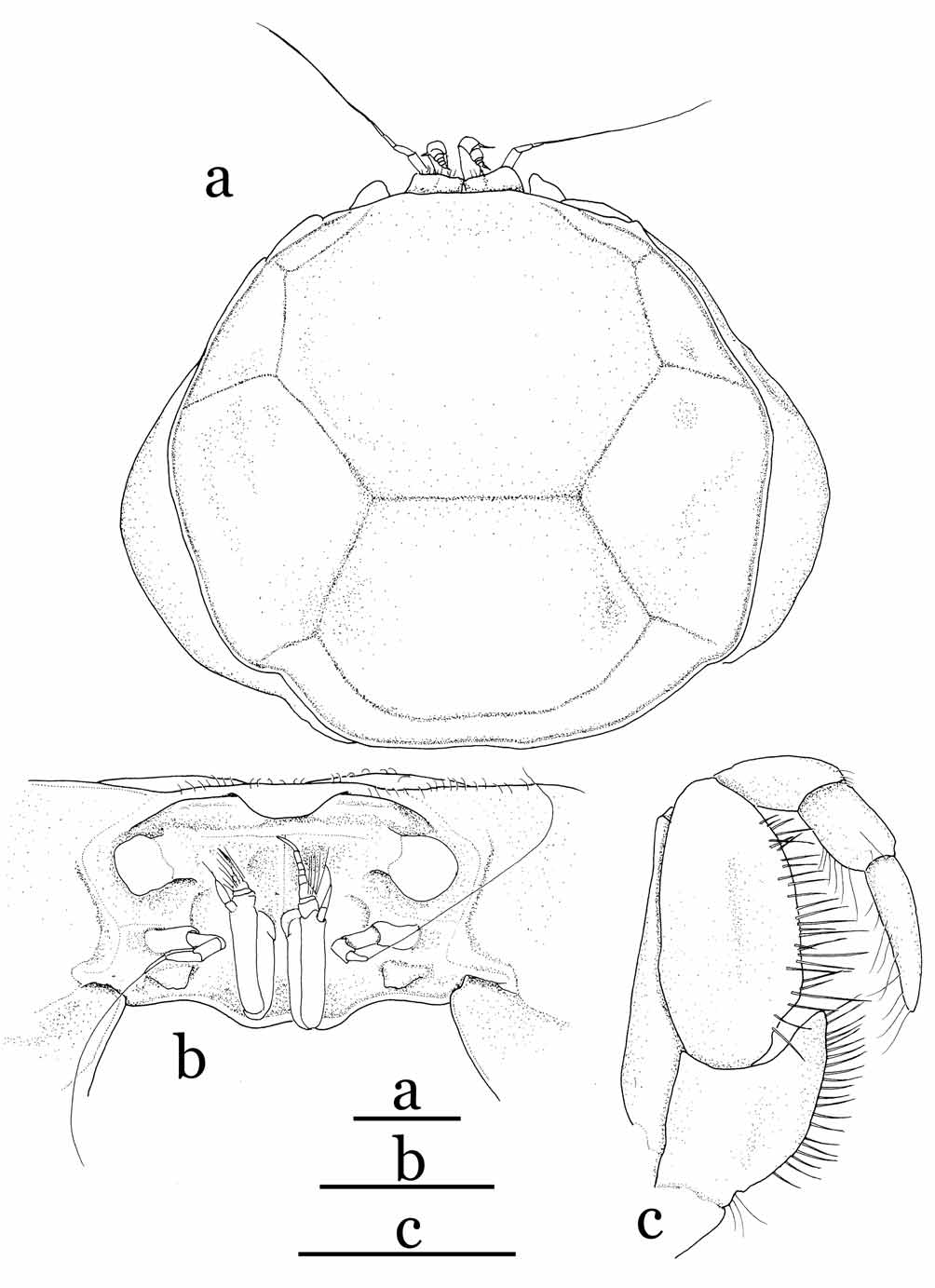
Diagnosis. Carapace subcircular, wider than long, dorsal surface flat, surrounded by continuous rim, regions clearly separated by grooves; rostrum vestigial, unilobed, deflexed, lateral surface below posterolateral margin without tooth. Anterior surface of cephalothorax (proepistome to epistome, orbit) nearly vertical in lateral view; antennular fossa very shallow, medially separated by thin septum (proepistome); epistome relatively long, with posterior margin produced and concave medially.
Eye slightly visible from dorsal view, base of ocular peduncle fused to orbit, immovable, gradually narrowed distally; cornea without trace of black pigment. Antennule with swollen basal article, largely protruding from fossa. Antenna with thin basal article, base of basal article located at outer lower region at a distance with respect to base of ocular peduncle, flagellum long. Third maxillipeds narrow, forming wide gape when closed; ischium with narrow, long distal inner extension, merus elongated, with inner, outer margins subparallel.
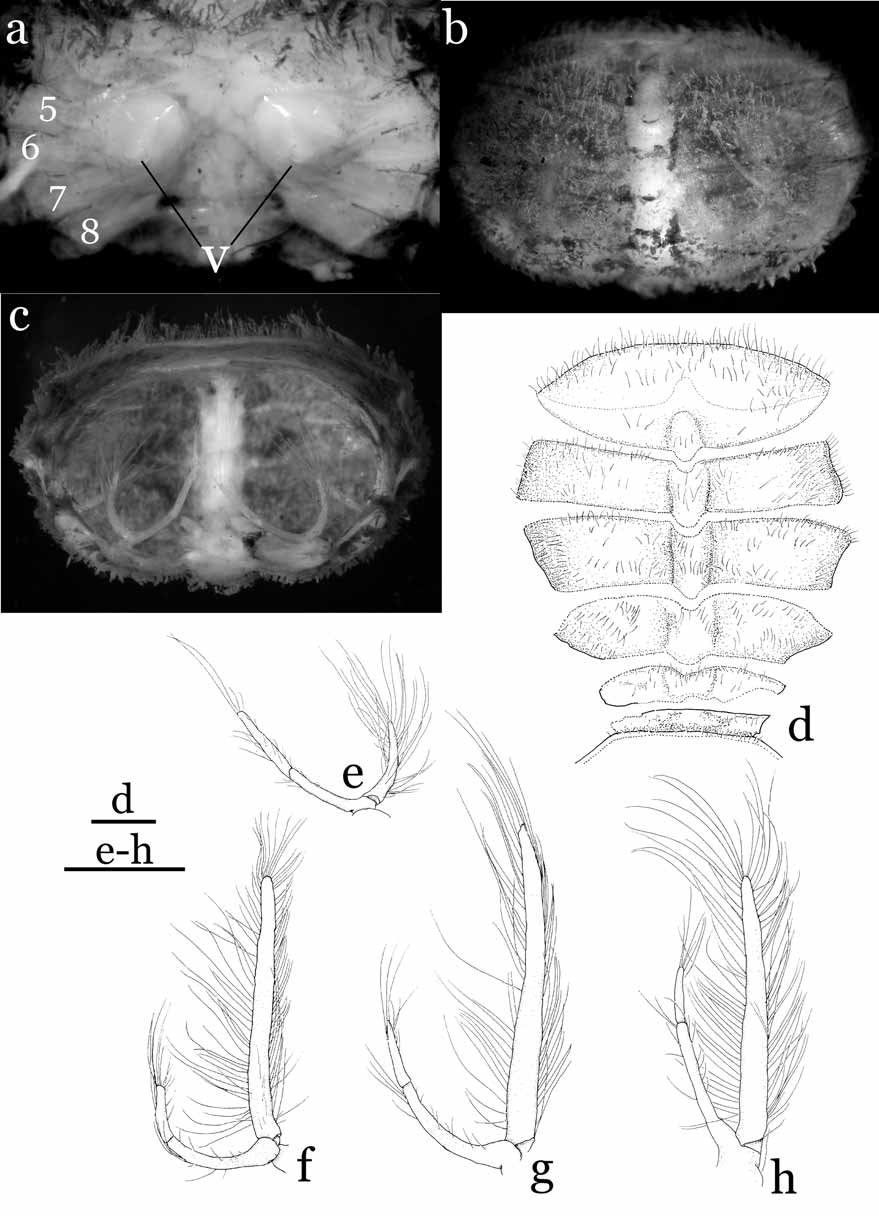
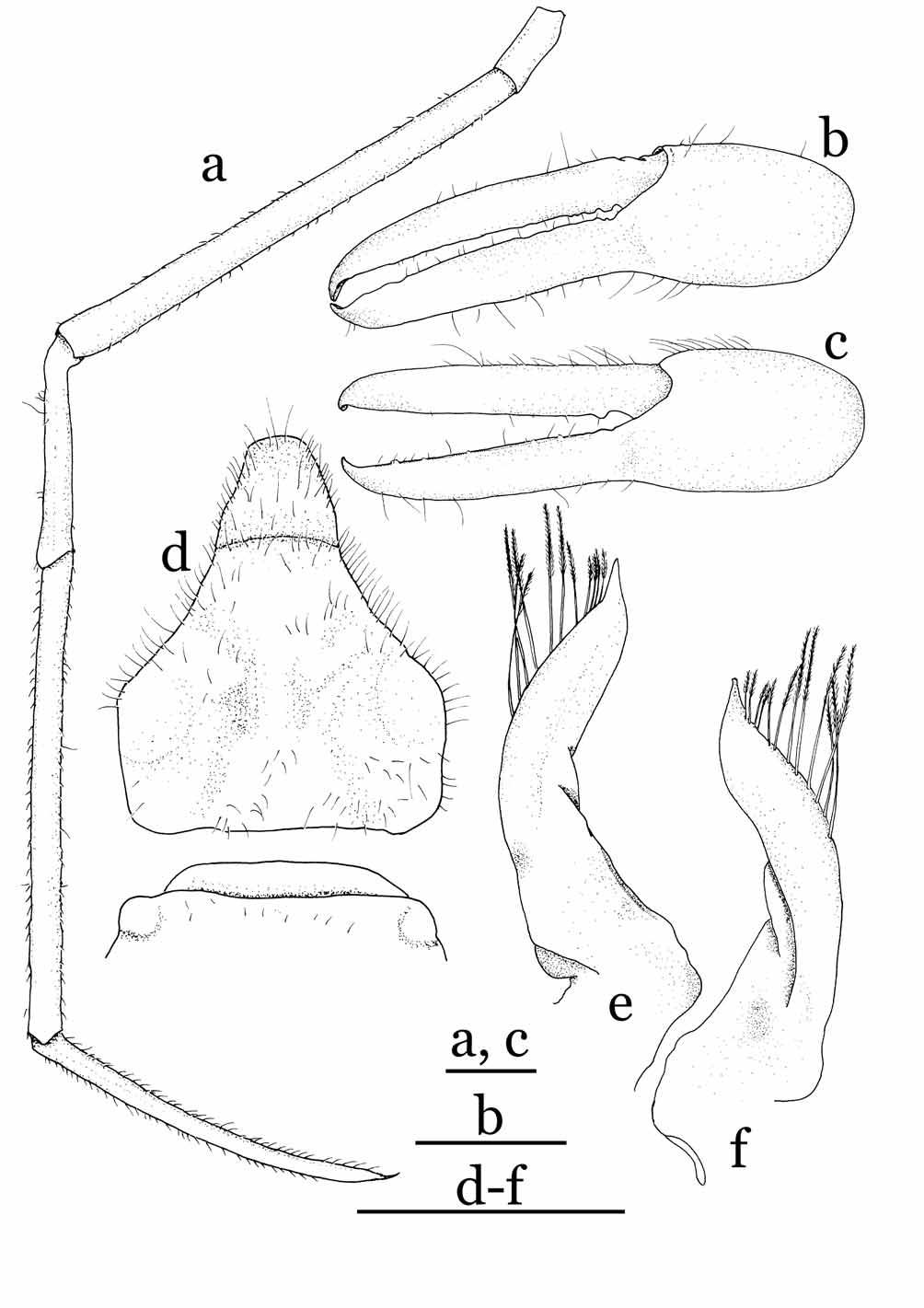
Male abdominal cavity narrow; sternal button for abdominal locking mechanism undiscernible. Female with wide cephalothorax cavity, margin of cavity connected to margin of abdomen-pleotelson by membrane, forming internal brood cavity; vulva of thoracic sternite 6 without basal mount along an imaginary line joining anterior margin of P2 coxae on medial fused plate of thoracic sternum. Chelipeds slender; chela with flat, blade-like fingers. Ambulatory legs extremely slender; dactyli slightly incurving, ending in sharp spine, without subterminal tooth. Penis located anterior to middle of sternite 8.
Male abdomen-pleotelson 4-segmented, third to fifth segments fused; first segment with protuberances on disto-lateral angles; fused segments narrowed in distal half, proximal half with subparallel lateral margins; pleotelson distinctly narrower than half width of abdomen. G1 strongly bent medially, tapering simply, dorsal side of inner margin lined with long, stiff setae; suture running from dorsal side of proximal part to outer margin of distal part; G2 with dilated base, proximal outer angle slightly visible posteriorly when abdomen closed. Female abdomen with an undivided plate formed by fusion of third segment to pleotelson; pleotelson only demarcated by shallow groove on outer side; second to fifth pleopods short, biramous, entirely enclosed inside abdominal brood cavity.
Etymology. The genus name, Sulaplax , is derived from the Sulawesi, and plax, “plate,” the suffix of the closely related genus Neorhynchoplax , alluding to their resemblance. Gender feminine.
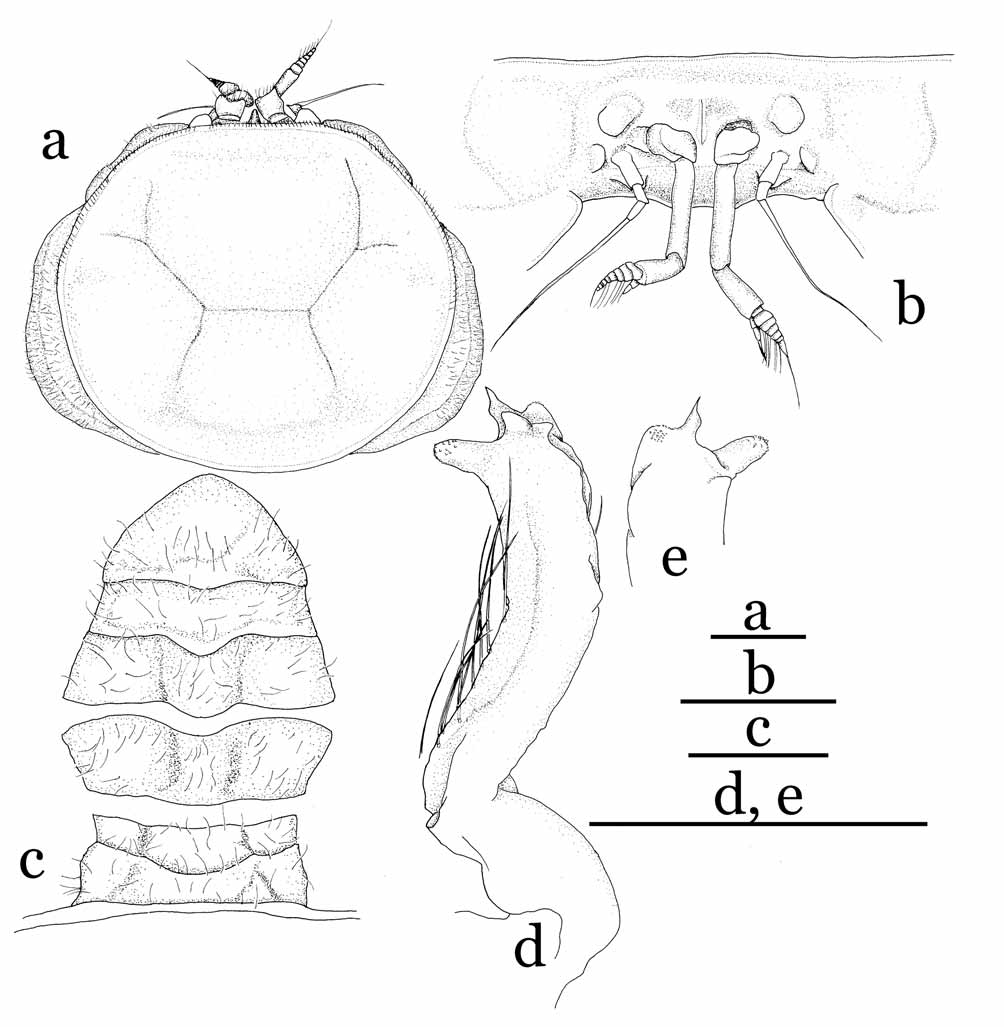
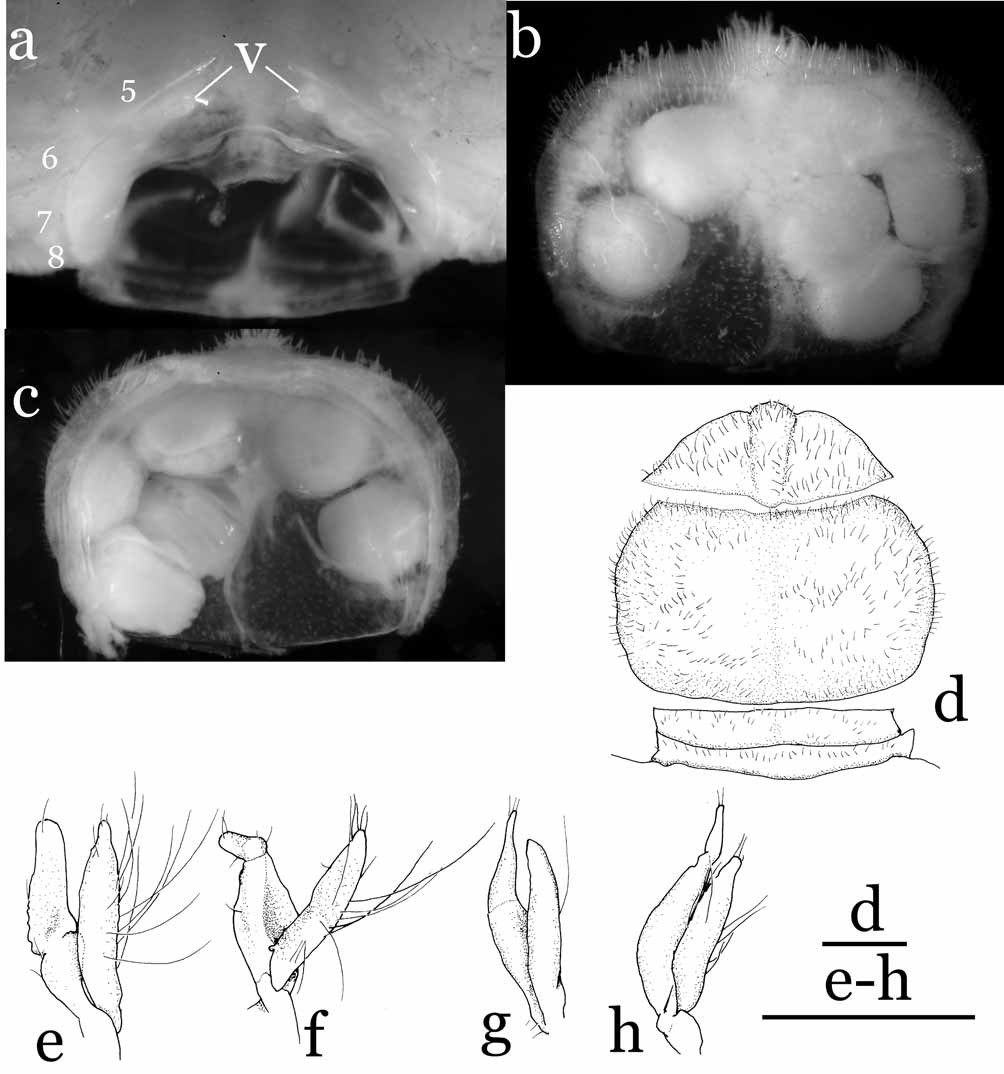
Remarks. Cavernicolous crabs often show varying degrees of characteristic morphological adaptations for a hypogeal lifestyle: long and slender ambulatory legs, depigmentation of the legs and carapace, and reduction of the cornea and eyes ( Rodríguez 1985; Guinot 1988; Ng 1991). Sulaplax gen. nov. is superficially similar to Cancrocaeca Ng, 1991 , in these cave-adapted characters. Sulaplax , however, differs significantly from Cancrocaeca in its relatively slender and gradually tapering G1 ( Fig. 5 View FIGURE 5 e, f), abruptly narrowed 4-segmented male abdomen with the third to fifth segments fused ( Fig. 5 View FIGURE 5 d), the female vulva lacking a basal mount ( Fig. 6 View FIGURE 6 a), and the short biramous female pleopods on the second to fifth abdominal segments ( Fig. 6 View FIGURE 6 e–h). In Cancrocaeca , the G1 is stout and with a complex apex ( Fig. 1 View FIGURE 1 d, e), the male abdomen is fully 6-segmented without fusion ( Fig. 1 View FIGURE 1 c), the female vulva bears a basal mount ( Fig. 2 View FIGURE 2 a), and the female pleopods are elongated and prominently setose on the second to fifth abdominal segments ( Fig. 2 View FIGURE 2 e–h). In addition, Sulaplax can be also distinguished from Cancrocaeca by the presence of a rostrum ( Figs. 4 View FIGURE 4 b) (absent in Cancrocaeca , Fig. 1 View FIGURE 1 a, b), well-defined dorsal carapace regions ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 a) (ill-defined in Cancrocaeca , Fig. 1 View FIGURE 1 a), and the absence of a longitudinal groove on the side walls of the carapace ( Figs. 4 View FIGURE 4 a) (present in Cancrocaeca , Fig. 1 View FIGURE 1 a) (see Table 1).
The brooding habits of Sulaplax species also differ remarkably from those of Cancrocaeca . The ovigerous females of C. xenomorpha hold the eggs only in the dome-shaped abdomen, which is marginally sheltered by the exopods of the third to fifth pleopods ( Fig. 2 View FIGURE 2 c, e–h). In contrast, the inner surface of the abdomen of S. ensifer spec. nov. is covered by a thin membrane and the reduced pleopods are present inside the membrane ( Fig. 6 View FIGURE 6 c, e–h). The membrane is connected to, and covers the margins of, the cephalothorax cavity and the eggs are thus kept inside the membrane of the abdomen and the cephalothorax cavities (equivalent to an internal brood cavity). Apart from the cave adaptation, the shape of the G1 and the fusion and shape of the male abdominal segments of Sulaplax suggest a close relationship to Neorhynchoplax , whose species dwell in a variety of intertidal to freshwater habitats in the Indo-West Pacific (see Lucas, 1980; Ng & Chuang, 1996). Furthermore, Sulaplax and some species of Neorhynchoplax possess an unusual internal brood cavity, which is otherwise known only in Crustaenia palawanensis ( Serène, 1971) and some species of Elamenopsis A. Milne Edwards, 1873 ( Lucas 1980: 191, fig. 8H; Lucas & Davie 1982: 406, 409; Ali et al. 1995: 3; Davie & Richer de Forges 1996: 261; Ng & Chuang 1996: 60, figs. 25, 26; Naruse et al. 2005: 5, fig. 1c–e; Naruse & Ng 2007a: 124). In contrast, the G1 and the male and female abdomens of Cancrocaeca are more similar to those of Limnopilos and Hymenicoides (see Remarks under Cancrocaeca and Table 1) than they are to those of Sulaplax . These relationships imply that troglobitic Sulaplax and Cancrocaeca are derived separately from two different ancestors; their superficial similarity is merely a remarkable example of convergence to a hypogeal lifestyle.
The characters which differentiate Sulaplax and Neorhynchoplax are mainly in the cephalothorax and its appendages. The anterior surface of the cephalothorax of Sulaplax is normal in that it appears flat in lateral view (dorso-anterior margin projecting beyond rest of cephalothorax in Neorhynchoplax ), the proepistome is relatively long and the epistome is relatively long and wide. The base of the antenna is located at outer lower region at a distance with respect to the base of the ocular peduncle ( Fig. 4 View FIGURE 4 b) (placed on just inner lower part of the base of the ocular peduncle in Neorhynchoplax ). The merus of the third maxilliped is elongated, and its inner and outer margins are subparallel (inner margin convex in Neorhynchoplax ). The dactylus of the third maxilliped is also elongated; its length is about twice the length of its propodus (dactylus clearly shorter than twice the length of the propodus in Neorhynchoplax ). Sulaplax can be also easily distinguished from Neorhynchoplax by its vestigial rostrum (always well developed in Neorhynchoplax ) and its reduced eyes (well developed in Neorhynchoplax ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |