
Ophryacus
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00748.x |
|
DOI |
https://doi.org/10.5281/zenodo.5619356 |
|
persistent identifier |
https://treatment.plazi.org/id/A67F87D0-FFAB-FFCC-FE86-FC2BFA9FFB97 |
|
treatment provided by |
Plazi |
|
scientific name |
Ophryacus |
| status |
|
OPHRYACUS COPE, 1887
Type species: Trigonocephalus [ Atropos ] Jan, 1859, by monotypy.
Etymology: The generic name is derived from the Greek ophrys, meaning brow, and the Latin acus, meaning pointed, obviously in reference to the distinctive supraocular spine-like scale.
Content: The genus Ophryacus contains only O. undulatus confined to the highlands of the Sierra Madre Oriental (Hidalgo, Veracruz, Puebla), the Mesa del Sur (Oaxaca), and the Sierra Madre del Sur (Oaxaca, Guerrero), where it occurs in pine-oak and cloud forest (Campbell & Lamar, 2004: map 84).
Common name: Mexican horned pitviper.
Definition and diagnosis: Rostral broader than high, moderately to distinctly concave; three preoculars, upper largest and undivided, middle not fused with supralacunal, lower small, somewhat excluded from margin of orbit; three to four supraoculars along dorsal margin of eye including supraocular spine; ten to 13 supralabials; lip margin not scalloped; nine to 14 infralabials; single scale above eye forming long, relatively slender spine, slightly compressed to subcircular in cross section, not occupying most of dorsal margin of orbit, tip pointed; adjacent scales along dorsal ocular margin often also modified, projecting slightly; canthals and internasals often raised into short spines or with especially high keels; scales in the supraocular region small and keeled; ten to 20 (usually 12–18) intersupraoculars; top of head covered with small scales, most having tubercular keels; second supralabial usually separated from prelacunal by single small subfoveal; subocular and supralabial series separated by two to four rows of small, roundish scales; 21 mid-dorsal scale rows; middorsals at midbody not noticeably broad, obtusely rounded; keel generally extending to tip of scale or nearly so, apical pits not apparent; free portion of apex of dorsal scales moderate in extent, barely overlapping contiguous scale; interstitial epidermal fold at cranial end of scale well developed; 157–178 ventrals; 37–57 subcaudals, divided; tail spine straight, about as long as preceding two to three subcaudals, pointed or obtusely rounded.
Frontal bones with concave dorsal surface, strongly elevated margins, moderately longer than wide; postfrontals moderate in size, not contacting frontal, comprising about equal amount of dorsal perimeter of orbit as parietals; posterolateral edges of dorsal surface of parietals forming distinct flat shelf continuing onto parietal as a raised ridge; junction between parietal and pro-otic irregular, not particularly angular; anterior portion of ectopterygoid possessing a shallow depression on medial side accommodating attachment of ectopterygoid retractor muscle; ectopterygoid noticeably longer than expanded, flattened base of pterygoid (posterior to articulation with ectopterygoid) with flat shaft tapering posteriorly; apex of choanal process positioned at about midlength on palatine, process moderately reduced in height, apex broadly rounded; dorsal surface of parietal roughly triangular; zero to one (usually zero) palatine teeth, seven to ten pterygoid teeth, seven to nine dentary teeth; pterygoid teeth extending to level of articulation of pterygoid with ectopterygoid; maxillary fang relatively short, only slightly longer than height of maxilla; fang at rest extending to level of suture between supralabials 7 and 8; splenial and angular bones fused; haemapophyses in contact distally.
Dorsum with zig-zag pattern; ground colour olivebrown, green, or grey, sometimes orange or yellow pigment present; dorsal scales usually finely mottled or speckled with black.
NATURAL HISTORY OF M. BARBOURI AND M. BROWNI
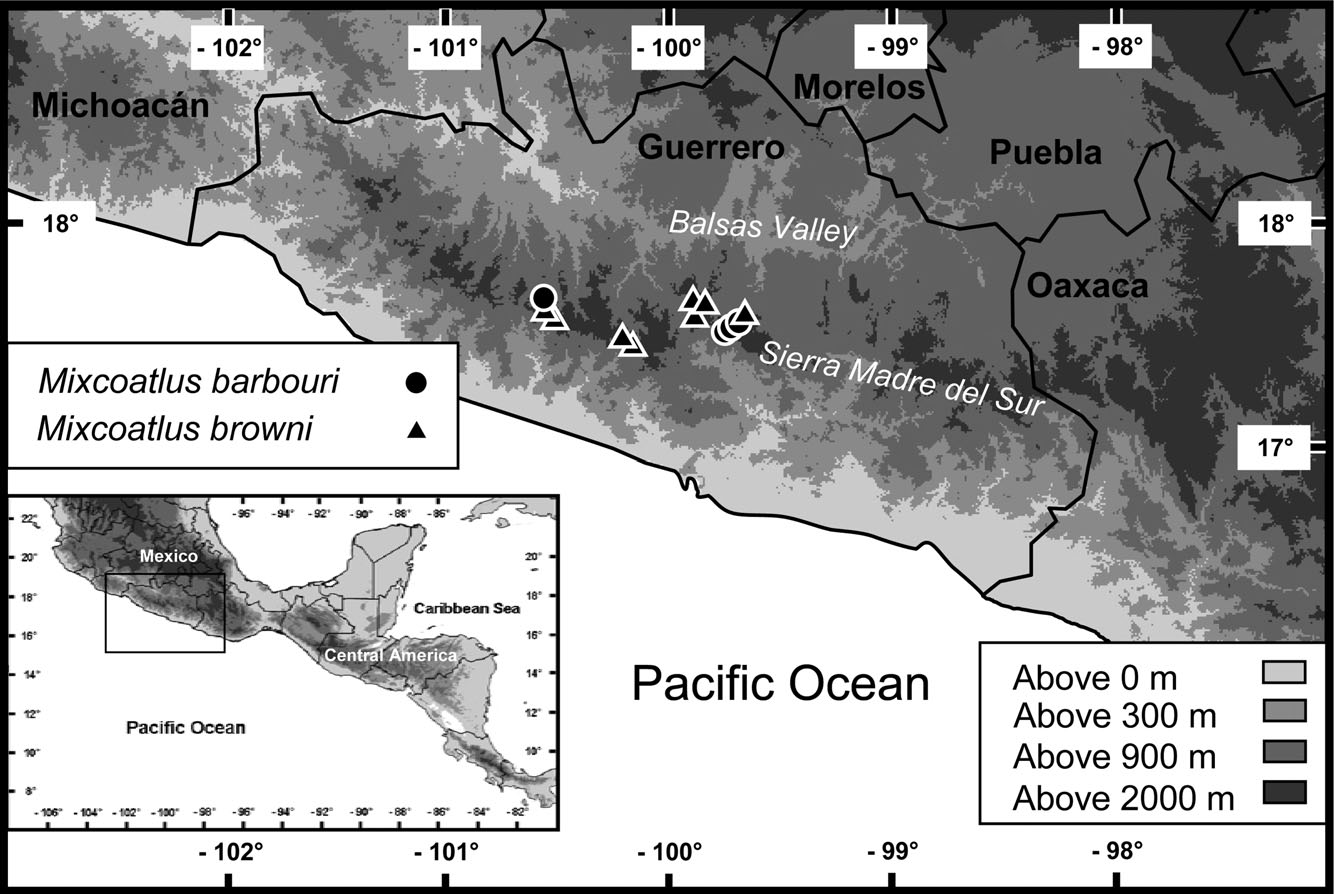
Similar to Ophryacus , Mixcoatlus is a pitviper genus endemic to the highlands of southern Mexico. Mixcoatlus appears to be found only in the western portion of the Sierra Madre del Sur of Guerrero ( M. barbouri and M. browni ) and north-western Oaxaca and south-eastern Puebla ( M. melanurus ), making it the most restricted genus of New World pitviper ( Fig. 6 View Figure 6 and map 83 in Campbell & Lamar, 2004, respectively). Mixcoatlus barbouri and M. browni are probably sympatric throughout much of their ranges, possess the same type locality (Omilteme, Guerrero), and have been found near each other in the western part of their ranges. Additionally, Crotalus intermedius omiltemanus , C. ravus , and O. undulatus have also been found near Omilteme and other highland areas in Guerrero, making their sympatry with M. barbouri and M. browni likely (Davis & Dixon, 1959; Campbell & Lamar, 2004). The highest confirmed elevation records for M. barbouri and M. browni are 2608 m (MZFC 21432) and 3296 m (KU 182762), respectively.
Campbell (1988) provides a detailed description of M. barbouri and M. browni habitat in the Sierra Madre del Sur and states that the higher elevations are dominated by pine-oak forest and cloud forest. Although it has been suggested that M. barbouri and M. browni inhabited cloud forest almost exclusively, observations of this species at lower elevations (Davis & Dixon, 1959; Campbell, 1988; this study) suggest that these species also occurs in upper pine-oak forest where it interdigitates with cloud forest. Additionally, two individuals of M. browni , one found in fir-pineoak forest (UTA R-4450) and the other ‘in bunchgrass on the sparsely wooded southern slope of Cerro Teotepec (KU R-182762)’, suggests that this species ‘at least inhabits several recognizable vegetation associations’ (Campbell, 1988: 8). Campbell (1988) also provides an investigation of the locality records for most of the specimens of M. barbouri and M. browni .
Both M. barbouri and M. browni are diurnal and are usually found basking, under cover, or moving during the day. While photographing a live M. browni , R. C. J. and E. N. S. observed an individual wrapping its tail around the hook to prevent falling (e.g. Fig. 4 View Figure 4 ). This same behaviour was also observed in the field in two other individuals. Although its tail is more prehensile than such terrestrial genera as Cerrophidion and Atropoides , we have no compelling evidence that M. browni is highly arboreal. It does ascend into low vegetation; J. A. C. observed one specimen coiled on top of a stump about 1.5 m above the ground and another in a low, woody shrub about 1.0 m high.
Few M. barbouri and M. browni have been kept in captivity. One specimen of M. barbouri (UTA R-15558) was kept for more than ten years (Campbell, 1988). An adult M. barbouri (MZFC 21432) and two M. browni (MZFC 21431 & UTA R-56265) were kept in captivity for several months. The M. barbouri was quite active and readily ate domestic white mice, whereas one adult (MZFC 21431) and one juvenile (UTA R-56265) M. browni required force feeding (A. Carbajal, pers. comm.). Specimens and scats of both M. barbouri and M. browni have contained rodent hair as well as the lizard Mesaspis gadovii (Campbell, 1988) . Mesaspis gadovii probably constitutes a large portion of the diet for these two pitvipers because of the great abundance of this lizard (Campbell, 1988). Two specimens of M. browni (MZFC 21431 & UTA R-56264) were captured within a few metres of Me. gadovii . Although only a few diet items have been identified, the diet of M. barbouri and M. browni probably includes lizards, orthopterans, and mammals, similar to that of Cerrophidion species, with less of their diet consisting of birds and amphibians (Campbell, 1988; Campbell & Solórzano, 1992; Campbell & Lamar, 2004; Jadin, 2007, 2010). Sceloporus adleri is abundant at these high elevations, representing another potential prey item. Many specimens of Thorius and Pseudoeurycea were collected in the vicinity of M. browni . A plethodontid salamander was found in the stomach of Cerrophidion petlalcalensis (López-Luna, Vogt & de la Torre-Loranca, 1999) and M. barbouri and M. browni may also consume them.
Of the 32 specimens of M. [ Agkistrodon ] browni and M. [ Cerrophidion ] barbouri examined in this study, 15 were M. barbouri and the other 17 specimens allocatable to M. browni (see Appendix S1 for details). We are aware of only three museum specimens not examined by us: MZFC 2880 and 2882 and a recently collected M. browni (field number JAC 27714).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |