
Leptomithrax lowryi, Forges & Ng, 2023
|
publication ID |
https://doi.org/10.3853/j.2201-4349.75.2023.1891 |
|
DOI |
https://doi.org/10.5281/zenodo.10957407 |
|
persistent identifier |
https://treatment.plazi.org/id/AC3B87B5-571F-FFD5-D8F3-FB9E261CA39B |
|
treatment provided by |
Felipe |
|
scientific name |
Leptomithrax lowryi |
| status |
sp. nov. |
Leptomithrax lowryi View in CoL sp. nov.
urn:lsid:zoobank.org:act:
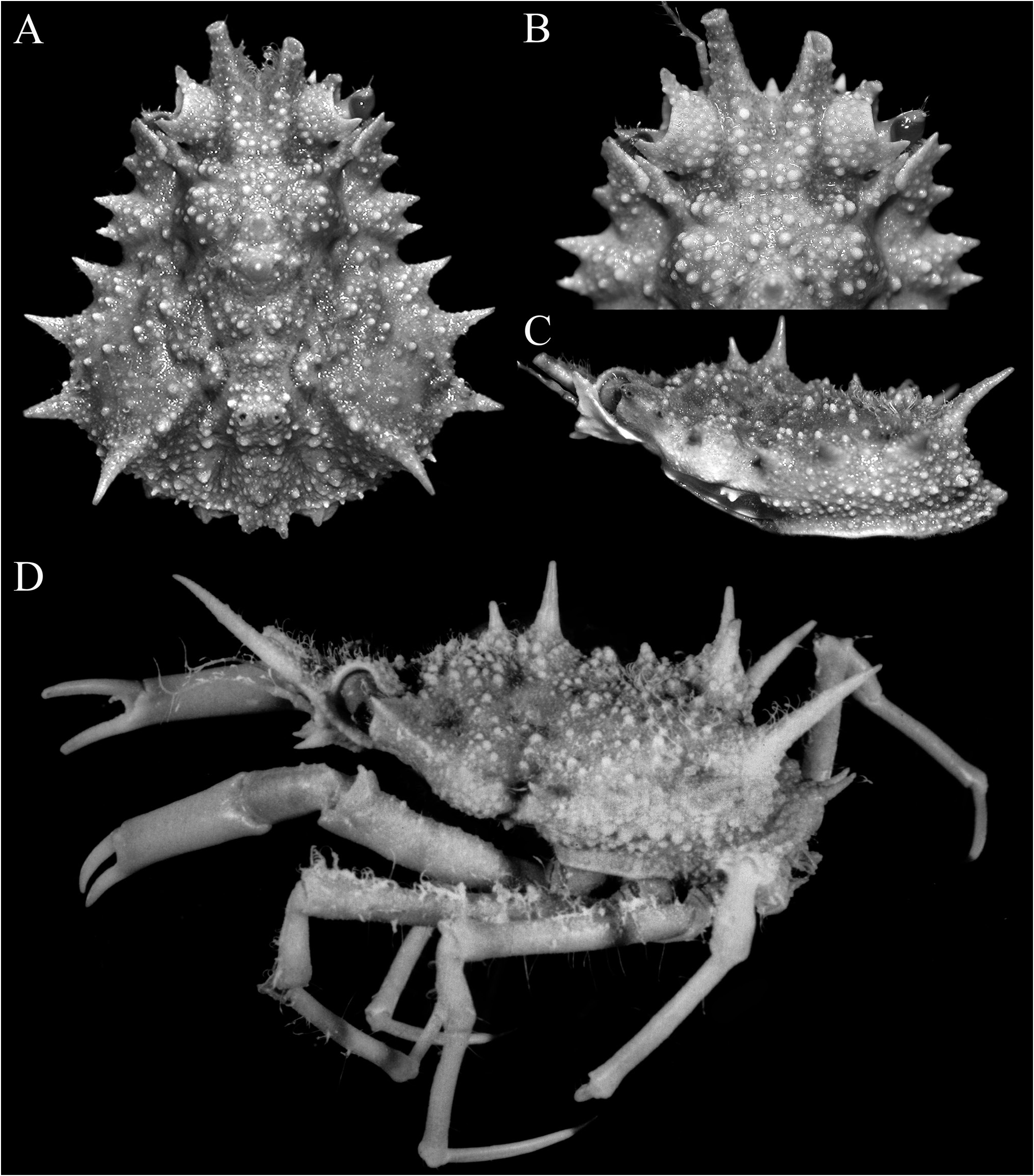
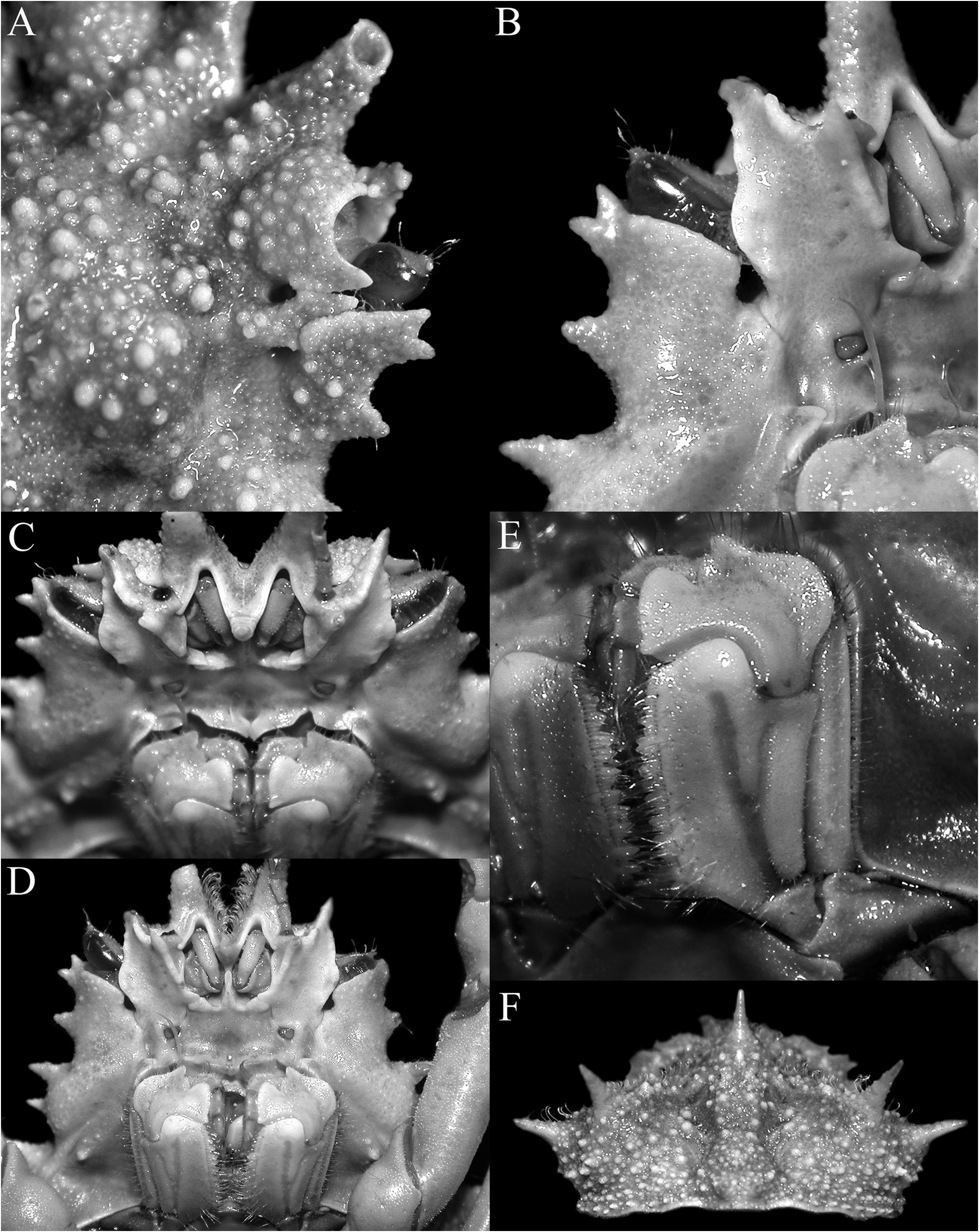
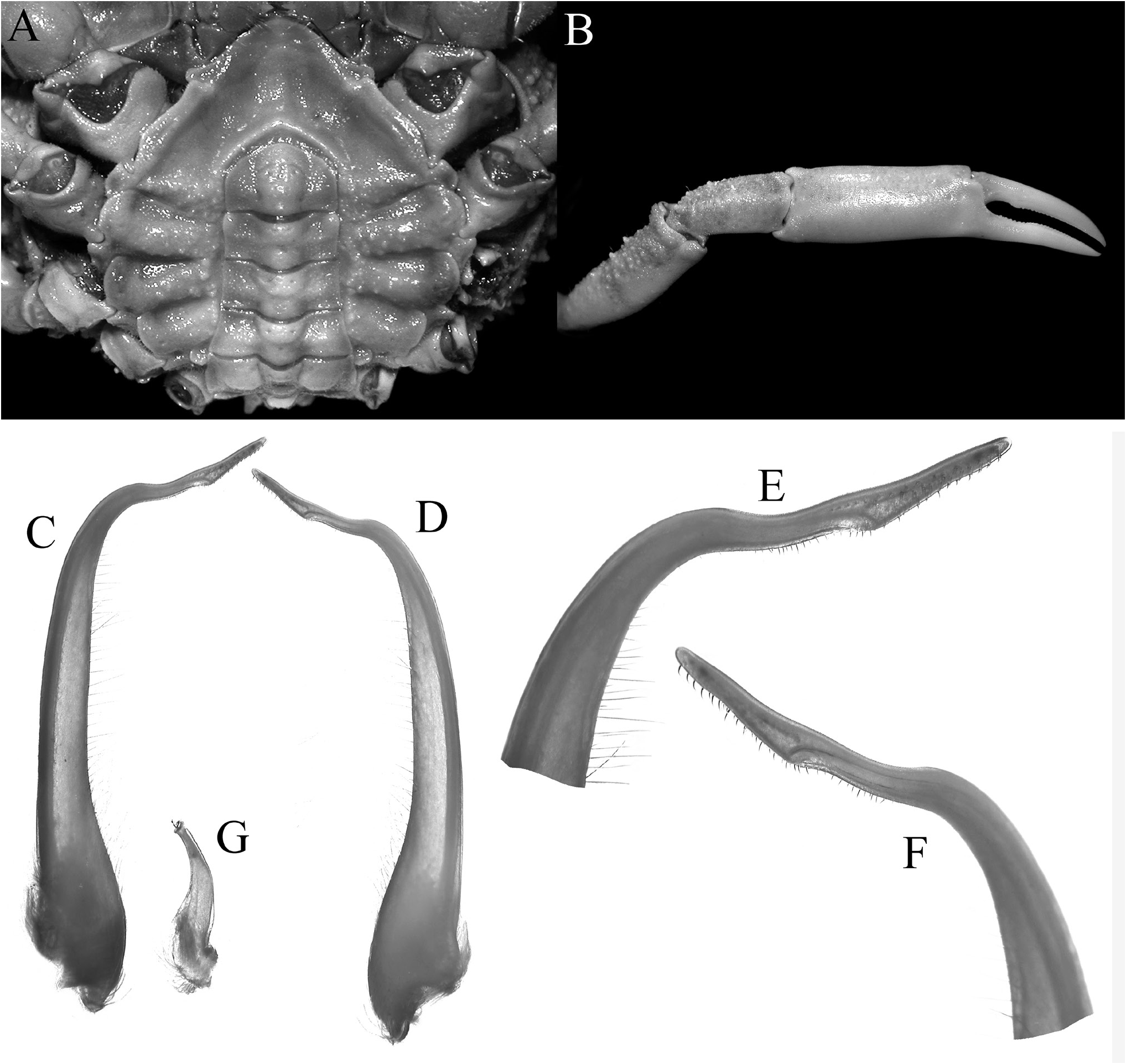
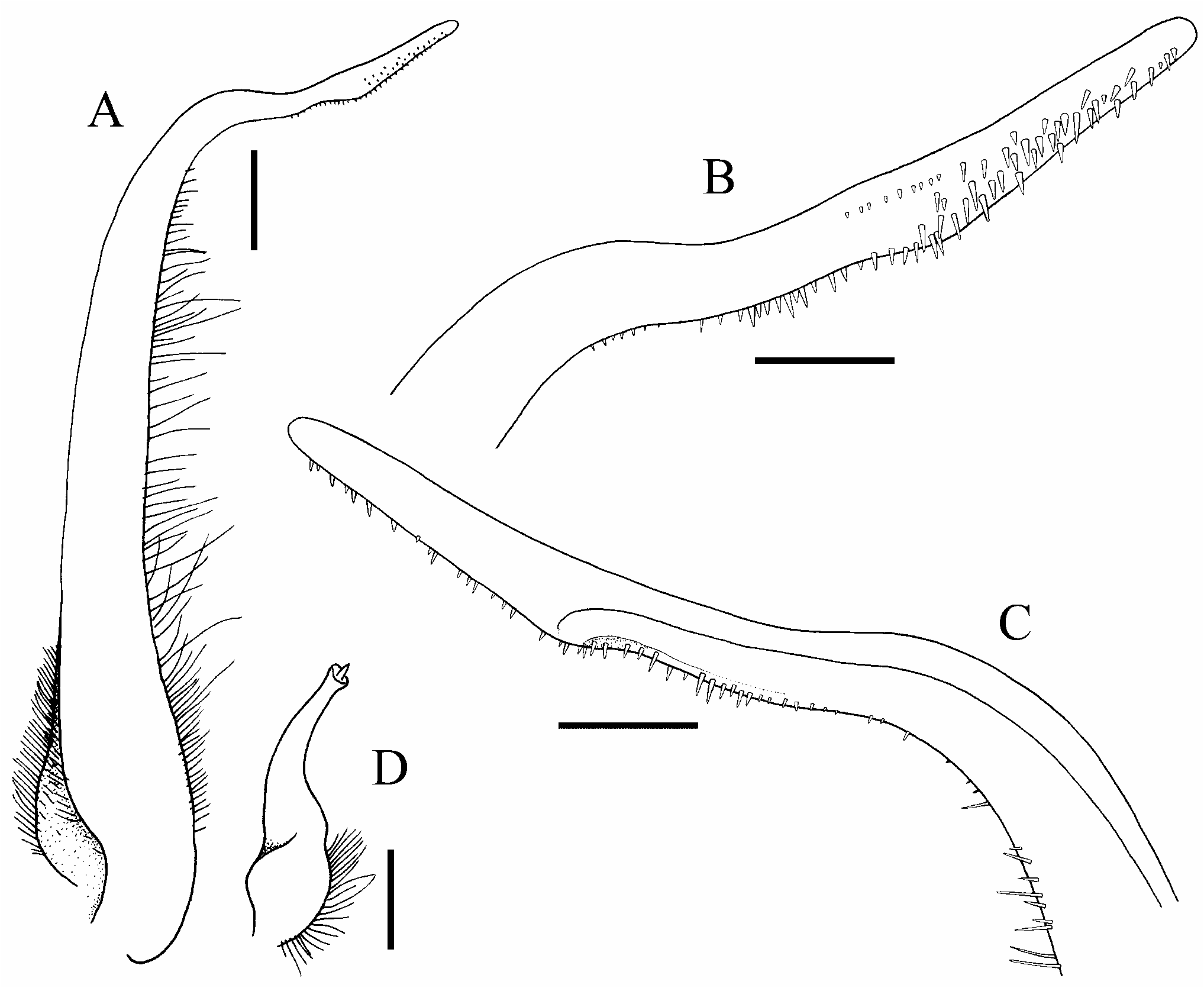
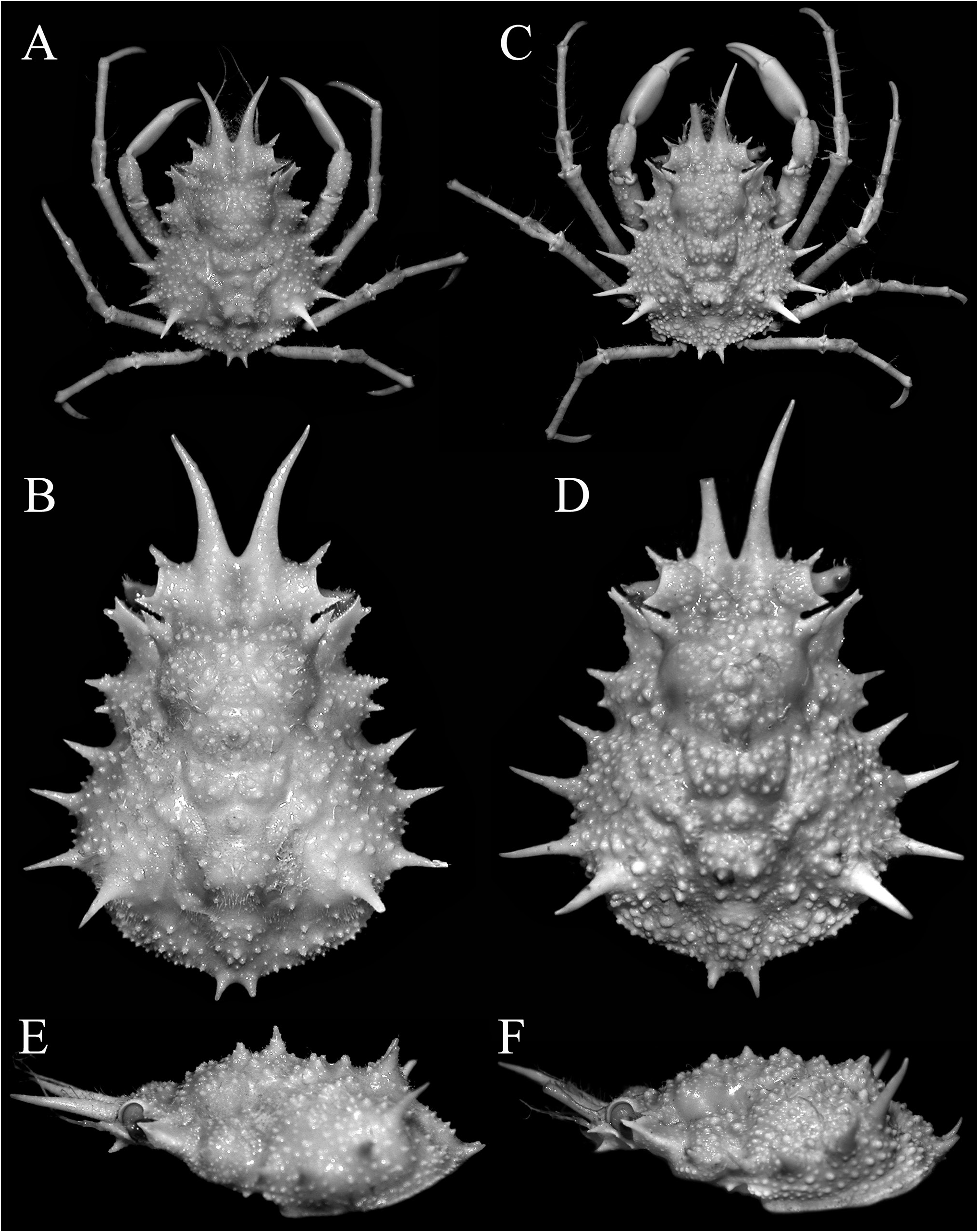
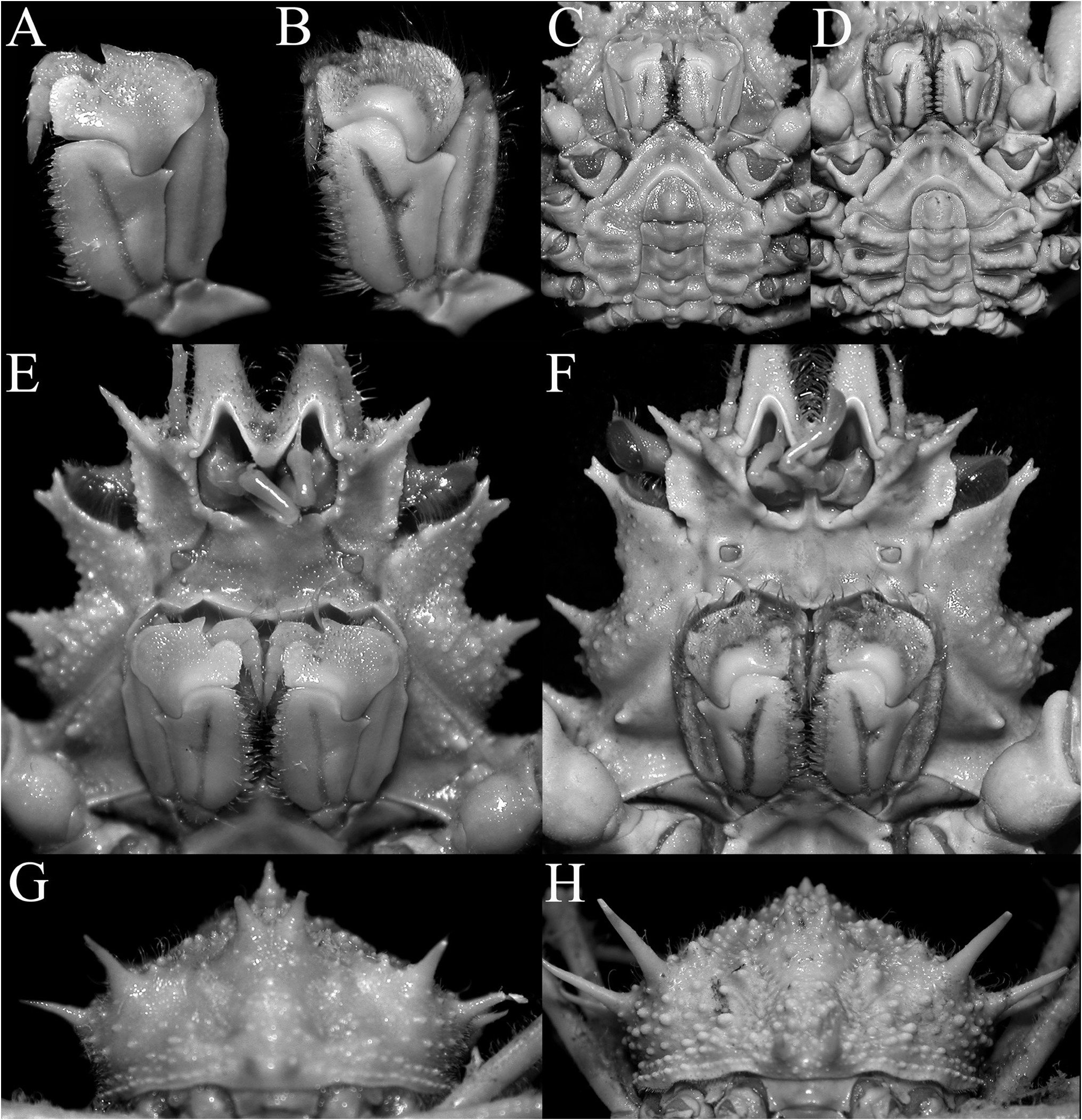
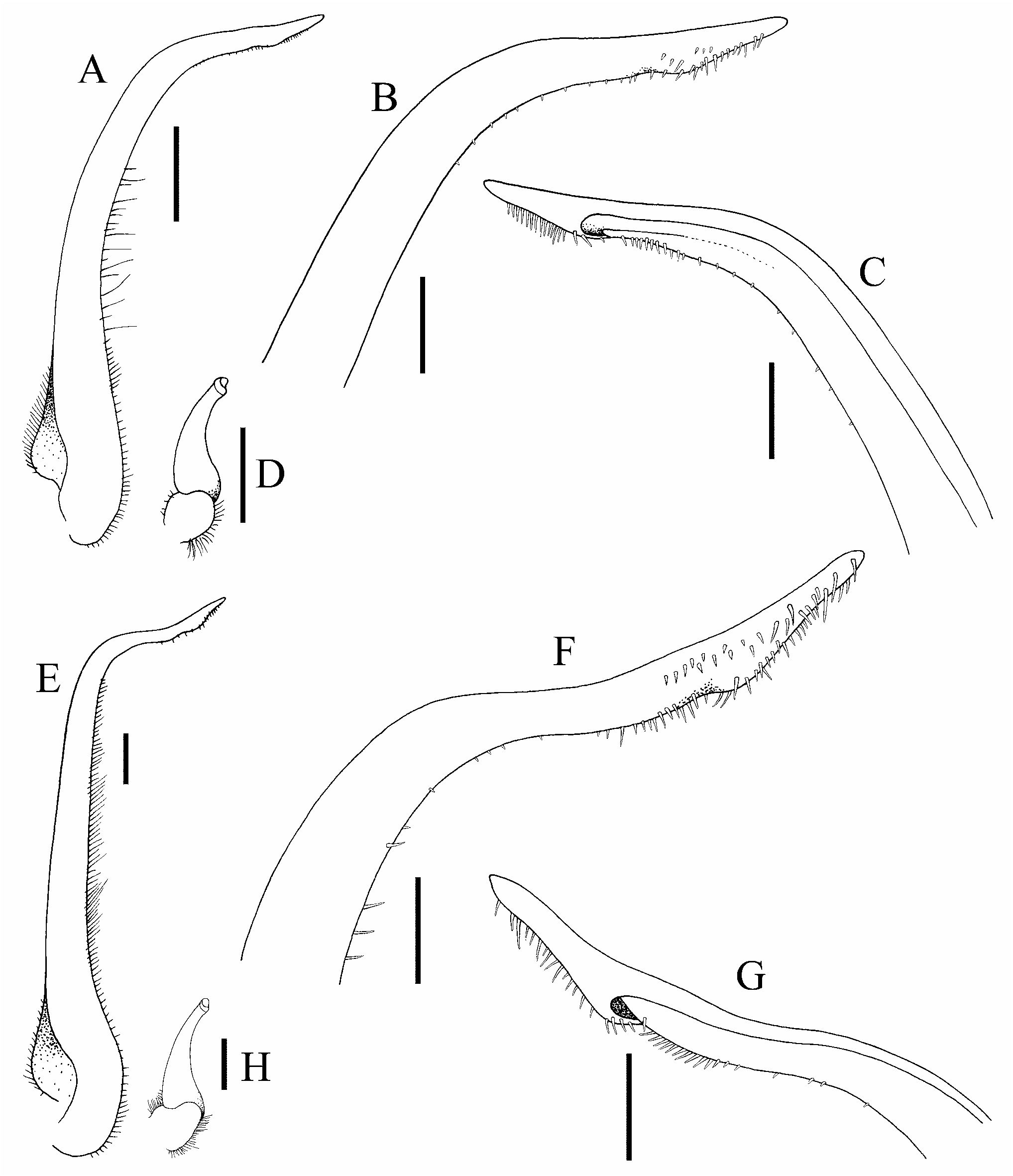
Figs 1–5 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5
Holotype: MNHN-IU-2022-187, male (cl 40.7 mm, pcl 34.3 mm, cw 37.0 mm, pcw 30.7 mm), station DW 5238, east of Atoll de la Surprise , New Caledonia, 18°06.5'S 163°03.4'E, 254–274 m, coll. SPANBIOS cruise, N.O. “Alis”, 22 July 2021 GoogleMaps . Paratypes: MNHN-IU-2018-5013, 1 male (cl 35.6 mm, pcl 29.3 mm, cw 29.4 mm, pcw 24.2 mm), 1 subadult female (cl 17.1 mm, pcl 14.8 mm, cw 13.7 mm, pcw 12.1 mm) same data as holotype; GoogleMaps MNHN-IU-2020-3639, 1 male (cl 28.0 mm, pcl 22.4 mm, cw 23.6 mm, pcw 17.7 mm), station DW 5171, south of Atoll Pelotas , New Caledonia, 19°01.4'S 163°26.8'E, 262–272 m, coll. SPANBIOS cruise, N.O. “Alis”, 8 July 2021; GoogleMaps ZRC 2022.0045 View Materials , 1 male (cl 32.3 mm, pcl 31.7 mm, cw 32.2 mm, pcw 27.5 mm), station DW 5207, east of Atoll de la Surprise , New Caledonia, 18°09'S 163°05.3'E, 239–248 m, coll. SPANBIOS cruise, N.O. “Alis”, 17 July 2021; GoogleMaps ZRC 2022.0046 View Materials , 1 female (cl 38.0 mm, pcl 30.8 mm, cw 30.9 mm, pcw 26.3 mm), 1 female (cl 38.1 mm, pcl 32.1 mm, cw 32.1 mm, pcw 26.2 mm), station DW 5207, east of Atoll de la Surprise , New Caledonia, 18°09'S 163°05.3'E, 239–248 m, coll. SPANBIOS cruise, N.O. “Alis”, 17 July 2021 GoogleMaps ; MNHN-IU-2021-3746, 1 female (cl 30.1 mm, pcl 24.5 mm, cw 26.4 mm, pcw 20.2 mm), 1 juvenile, station DW5174, south of Atoll Pelotas , New Caledonia, 18°57.8'S 163°23.2'E, 320–342 m, coll. SPANBIOS cruise, N.O. “Alis”, 8 July 2021 GoogleMaps ; MNHN-IU-2022-186, 1 male (cl 30.8 mm, pcl 21.9 mm, cw 25.0 mm, pcw 16.6 mm), 1 subadult female (cl 14.1 mm, cw 10.8 mm), station DW727, slope southwest of Dumbéa passage, New Caledonia, 22°48.03'S 167°29.03'E, 299–302 m, coll. BATHUS 2 cruise, N.O. “Alis”, 12 May 1993 GoogleMaps .
Diagnosis. Carapace very wide posteriorly, inflated medially; dorsal surface strongly granulous, anterior half gently convex. Pseudorostral spines divergent. Supraocular eave wide; intercalated spine narrow, distally blunt, almost totally closing gap between antorbital and postorbital spines; postorbital spine long, foliaceous, divided distally into 2 spines, proximal angle with distinct tooth. Basal antennal article surface almost flat, outer margin distinctly convex. Third maxilliped with junction of merus and ischium gently swollen; ischium with distinctly Y-shaped median furrow, anteroexternal angle forming large tooth with rounded tip. Ambulatory legs, especially meri, with smooth dorsal margin. Male thoracic sternites with lateral surfaces shallowly but distinctly excavated. Male telson subpentagonal. G1 slender, distal one-third curving outwards; distal part relatively long, gently sinuous.
Description. Carapace pyriform, very wide posteriorly; dorsal surface very granulous, anterior half gently convex; gastric and branchial regions distinct, separated by distinct grooves; carapace and pereiopods covered with scattered short setae, not obscuring surface or margins ( Figs 2 View Figure 2 , 3A–C View Figure 3 ). Pseudorostral spines sharp, gently curved, diverging, cylindrical in cross-section, lateral margins without distinct spines or tubercles ( Figs 1 View Figure 1 , 2 View Figure 2 , 3A–C View Figure 3 ). Proepistome with sharp, gently curved spine, visible in dorsal view ( Figs 3B, C View Figure 3 , 4C View Figure 4 ). Supraocular eave wide, forming sharp anterior and posterior angles, deeply convex above eye; intercalated spine narrow, distal part acute with proximal part subtruncate, almost totally closing gap between antorbital and postorbital spines with narrow basal fossae visible; postorbital spine long, foliaceous, divided distally into 2 short spines, proximal angle with distinct tooth ( Figs 2 View Figure 2 , 3A, B View Figure 3 , 4A, B View Figure 4 ). Carapace inflated medially, with granulated tubercles but no spine; posterior margin of carapace with 2 short sharp divergent spines; hepatic area with 2 strong spines directed outward; 4 lateral branchial spines, with posterolateral part armed with 3 sharp long spines, posteriormost one subdorsal in position, directed posteriorly and upwards, distinctly longer than other spines ( Figs 2C View Figure 2 , 3D View Figure 3 , 4F View Figure 4 ). Basal antennal article wide, surface smooth, almost flat, completely fused with carapace, internal distal margin forming blunt tooth, outer margin distinctly convex, smooth; urinary article with subrectangular orifice; separated from postorbital spine by basal fossa ( Fig. 4B View Figure 4 ); antenna deeply protected, far from orbit, flagellum long ( Figs 3B, D View Figure 3 , 4A–D View Figure 4 ). Eye short, protected by inner surface of postorbital tooth; cornea ovate, large, with small granule at distal tip ( Fig. 4C–D View Figure 4 ). Epistome medially depressed with smooth surface ( Fig. 4D View Figure 4 ). Epistome transversely rectangular; posterior margin with broad, subtruncate median plate, with deep median fissure; separated from lateral parts by deep V-shaped cleft ( Fig. 4C, D View Figure 4 ).
Third maxilliped short, with smooth surface, bordered by setae on margin of merus; junction of merus and ischium gently swollen medially; ischium with distinctly Y-shaped median furrow, outer branch of furrow about three-quarters length of inner branch, anteroexternal angle with large tooth with rounded tip; merus short, anteroexternal angle auriculiform, distal median margin with rounded tooth, separated from inner part by deep cleft; exopod relatively slender, reaching to median part of merus, with long flagellum ( Fig. 4D, E View Figure 4 ).
Chelipeds not elongate; merus short, dorsal margin lined with rounded and sharp granules, without distal spine; carpus short, proximal dorsal surface with granules, rest of surface smooth; chela slender, surfaces smooth; fingers shorter than palm, almost straight, cutting margins with denticles; proximal part of cutting margin of dactylus with low concavity, followed by low, wide tooth ( Figs 2 View Figure 2 , 3D View Figure 3 , 5B View Figure 5 ).
Ambulatory legs, especially meri, proportionately long; first leg longest, fourth leg shortest; merus subcylindrical, dorsal margin smooth, distal angle of dorsal margin angular but not spiniform or dentiform; propodus longer than dactylus; dactylus gently curved with corneous tip ( Figs 2 View Figure 2 , 3D View Figure 3 ).
Thoracic sternites 1–4 completely fused; proximal part of sternite 4, median surface of sternites 5 and 6 with low granules; median longitudinal ridge on sternites 3 and 4 relatively low, with lateral surfaces shallowly but distinctly excavated; distal part of sternopleonal cavity demarcated by relatively sharp C-shaped ridge; part of sternite 8 exposed when male pleon closed ( Fig. 5A View Figure 5 ).
Male pleon subrectangular in shape, with 6 free somites and telson; telson subpentagonal with convex distal margin; somite 3 widest, reaching coxae of last ambulatory legs; somite 4 trapezoidal; somites 5 and 6 rectangular; ( Fig. 5A View Figure 5 ).
G1 slender, distal one-third curving outwards; distal part relatively long, gently sinuous, ventral margin lined with distinct spinules ( Figs 5C–F View Figure 5 , 6A–C View Figure 6 ). G2 short, ca. one-fifth length of G1; tip cup-like with very short flagellum ( Fig. 6D View Figure 6 ).
Colour. Dorsal surfaces orange ( Fig. 1 View Figure 1 ).
Etymology. This species is named in honour of our friend and colleague James (Jim) Kenneth Lowry from the Australian Museum. A prolific taxonomist of peracarids, especially amphipods of the superfamily Lysianassoidea , he was also responsible for an important revision of the giant deep-sea isopods of the genus Bathynomus A. Milne-Edwards, 1879 .
Remarks. Griffin & Tranter (1986: 208) divided Leptomithrax into several species groups, separated by various carapace and third maxilliped features. One group, with L. sternocostulatus and L. bifidus , has the distal part of the postorbital tooth truncate or bifid, the intercalated spine is mostly excluded from the rim of the incipient supraorbital margin, the junction of the ischium and merus of the third maxilliped is swollen, and the surfaces of the thoracic sternum and/or pleon are excavated. Griffin & Tranter (1986: 208) placed L. sinensis elsewhere as its third maxilliped and sternal conditions were not known, but Wong et al. (2018) showed it was close to L. bifidus and should also be in this group of species. Two other species, L. depressus and L. eldredgei also belong to this group.
Leptomithrax sternocostulatus can immediately be separated from other members of this group in that it only has three lateral branchial spines and the excavations on the male thoracic sternum and pleon are very deep with rims around the depressions; all the others have four lateral branchial teeth and the excavations on the ventral surface are distinctly shallower (cf. Grant & McCulloch, 1906: pl. 3 fig. 2; Poore, 2004: figs. 111b, c, 115p). Leptomithrax depressus is distinct in that the anterior one-third of the dorsal surface of the carapace is more flattened than any of the congeners and can also be separated from L. lowryi sp. nov. in having the male pseudorostral spines proportionately shorter and subparallel ( Richer de Forges, 1993: fig. 7a, b) (versus long and distinctly diverging in L. lowryi sp. nov.; Fig. 2 View Figure 2 ); the proximal angle of the postorbital tooth is demarcated by a low rounded lobe ( Richer de Forges, 1993: fig. 7b) (versus a distinctly dentiform lobe in L. lowryi sp. nov.; Fig. 3A, B View Figure 3 ); the intercalated spine is short, triangular with a rounded tip ( Richer de Forges, 1993: fig. 7b) (versus spine longer with a sharp distal part and a more truncate base in L. lowryi sp. nov.; Fig. 3A, B View Figure 3 ); the fourth subdorsal branchial spine is stouter and slightly longer than the third spine ( Richer de Forges, 1993: fig. 7b) (versus spine is distinctly longer and more slender in L. lowryi sp. nov.; Figs 2 View Figure 2 , 3A View Figure 3 ); the ambulatory legs, in particular the meri, are proportionately shorter ( Richer de Forges, 1993: fig. 7a) than in L. lowryi sp. nov. ( Fig. 2 View Figure 2 ); the surface of the merus of the ambulatory legs, especially upper surface, is coarsely granular and uneven ( Richer de Forges, 1993: fig. 7b) (versus smooth in L. lowryi sp. nov.; Fig. 2 View Figure 2 ); and the distal quarter of the G1 is gently curved ( Richer de Forges, 1993: fig. 7c) (versus distal part of the G1 is sinuous in L. lowryi sp. nov.; Figs 5C–F View Figure 5 , 6A–C View Figure 6 ).
Leptomithrax eldredgei can be separated from L. lowryi sp. nov. by the more slender carapace and the proportionally shorter lateral branchial spines (compare Richer de Forges & Ng, 2015: fig. 1A with Figs 2 View Figure 2 , 3A View Figure 3 ); the male pseudorostral spines are proportionately shorter and subparallel ( Richer de Forges & Ng, 2015: fig. 1A, D) (versus long and distinctly diverging in L. lowryi sp. nov.; Fig. 2 View Figure 2 ); the postorbital tooth appears foliaceous and almost entire, with the distal angle marked by a low bifurcation and the posterior angle broad and rounded ( Richer de Forges & Ng, 2015: fig. 1A) (versus distal and proximal parts separated by deep cleft, the proximal angle marked by a distinct tooth in L. lowryi sp. nov.; Fig. 3A, B View Figure 3 ); the submedian sulcus on ischium of third maxilliped is almost entire and the anteroexternal angle has a sharp tooth ( Richer de Forges & Ng, 2015: fig. 1C) (versus the sulcus on the ischium is Y-shaped with the anteroexternal angle rounded in L. lowryi sp. nov.; Fig. 4E View Figure 4 ); and the distal part of the G1 is gently curved and proportionately shorter ( Richer de Forges & Ng, 2015: fig. 2) (versus distal part of the G1 is sinuous in L. lowryi sp. nov.; Figs 5C–F View Figure 5 , 6A–C View Figure 6 ).
The morphology of Leptomithrax lowryi sp. nov. appears to be closest to two east Asian species, L. bifidus Ortmann, 1893 and L. sinensis Rathbun, 1916 ( Figs 7–9 View Figure 7 View Figure 8 View Figure 9 ). The identity of L. sinensis was clarified by Richer de Forges & Ng (2015) from the type which is a dried carapace; with Wong et al. (2018) redescribing the species from fresh material from Taiwan and Japan and comparing it at length with L. bifidus . Leptomithrax lowryi sp. nov. differs from the two species in that its carapace is proportionately wider posteriorly ( Figs 2 View Figure 2 , 3A View Figure 3 ) than in L. bifidus and L. sinensis ( Fig. 7A–D View Figure 7 ); the posterolateral spines are relatively stout ( Fig. 3A View Figure 3 ) (versus more slender in L. bifidus and L. sinensis , Fig. 7B, D View Figure 7 ); the intercalated spine is blunt ( Fig. 3A, B View Figure 3 ) (versus sharp in L. bifidus and L. sinensis , Fig. 8E, F View Figure 8 ); the pseudorostral spines are less curved and diverging ( Figs 2 View Figure 2 , 3A View Figure 3 ) (versus more curved and clearly diverging in L. bifidus and L. sinensis , Fig. 7B, D View Figure 7 ); the deep furrow on the ischium of the third maxilliped is distinctly Y-shaped with the outer branch about three-quarters the length of the inner one ( Fig. 4D, E View Figure 4 ) (versus furrow weakly Y-shaped with the outer branch less than half the length of the inner one in L. sinensis and L. bifidus , Fig. 8A, B, E, F View Figure 8 ); the surface of the basal antennal article is almost flat ( Fig. 4B–D View Figure 4 ) (versus gently depressed in L. sinensis and L. bifidus , Fig. 8E, F View Figure 8 ); the outer margin of the basal antennal article is distinctly convex and smooth ( Fig. 4B–D View Figure 4 ) (versus almost straight, gently convex and may be lined with granules in L. sinensis and L. bifidus , Fig. 7E, F View Figure 7 ); and the median longitudinal ridge on fused male thoracic sternites 3 and 4 is relatively low, with the lateral surfaces gently depressed ( Fig. 5A View Figure 5 ) (versus with median longitudinal ridge high, separating the two deep lateral depressions in L. sinensis and L. bifidus , Fig. 8C, D View Figure 8 ). The telson of the male of L. lowryi sp. nov. resembles that of L. sinensis , being subquadrate-subpentagonal in shape ( Figs 5A View Figure 5 , 8D View Figure 8 ), whereas that of L. bifidus is distinctly more triangular ( Fig. 8C View Figure 8 ). The G1 of L. lowryi sp. nov. is distinctly different from that of L. bifidus in that the distal part is sinuous rather than gently curved ( Figs 5C–F View Figure 5 , 6A–C View Figure 6 versus Fig. 9A–C View Figure 9 ). The G1 of L. lowryi sp. nov. is most similar to L. sinensis except that the distal spinous part is proportionately longer in the former species ( Fig. 6A–C View Figure 6 versus Fig. 9E–G View Figure 9 ).
It is noteworthy that L. lowryi sp. nov. from New Caledonia is morphologically closest to two East Asian species, L. bifidus and L. sinensis , rather than to taxa from Australia, viz. L. tuberculatus , L. sternomaculatus and L. depressus (cf. Griffin, 1966; Richer de Forges, 1993; Poore et al., 2008).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
SuperFamily |
Majoidea |
|
Family |
|
|
SubFamily |
Majinae |
|
Genus |