
Hyperoche medusarum ( Kröyer, 1838 ), Kroyer, 1838
|
publication ID |
https://doi.org/10.11646/zootaxa.3905.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:A47AE95B-99CA-42F0-979F-1CAAD1C3B191 |
|
DOI |
https://doi.org/10.5281/zenodo.6114486 |
|
persistent identifier |
https://treatment.plazi.org/id/AE418800-FFD7-FF82-FF3F-FDE062BDFDEA |
|
treatment provided by |
Plazi |
|
scientific name |
Hyperoche medusarum ( Kröyer, 1838 ) |
| status |
|
Hyperoche medusarum ( Kröyer, 1838) View in CoL
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Metoecus medusarum Kröyer, 1838: 288 –291; pl. 3, fig. 15a-n.— Milne-Edwards 1840: 78. White 1847: 91. Gosse 1853: 367. White 1857: 207. Boeck 1871: 86.
Metoicus medusarum — Kinahan 1859: 83.
non— Hyperia medusarum View in CoL — Bate 1862: 295–296; pl. 49, fig. 1, (= Hyperia View in CoL sp.).
Hyperia medusarum View in CoL —Goës 1865: 534.
Tauria medusarum — Sars 1882: 75.
Hyperoche medusarum View in CoL — Hansen 1888: 58 –59. Stebbing 1888: 1399. Stappers 1911: 77. Stephensen 1912: 557 & 609 (lists), 612. Stephensen 1913b: 99 –100. Stephensen 1923a: 13 –15, chart 1. Stephensen 1923b: 7. Stephensen 1924: 79. Schellenberg 1927: 630 –631, fig. 38. Stephensen 1928: 30 –31, figs. 4.7–4.8. Pirlot 1929: 118 –119. Stephensen 1929: 42, fig. 13.3. Barnard 1930: 415 (key). Stephensen 1932: 375 (list). Stephensen 1933: 61. Behning 1939: 361 –362. Pirlot 1939: 38. Stephensen 1940: 5, 84 & 98 (tables). Stephensen 1942: 458, 504 (table). Stephensen 1944: 9, tables (pp. 145, 149 & 160). Bousfield 1951: 139, fig. 3 (map), passim. Dunbar 1954: 782. Bulycheva 1955: 1043 (table). Hurley 1956: 14. Vinogradov 1956: 211. Irie 1957: 10 (table). Irie 1958: 107 (table). Irie 1959: Table 4. Grice & Hart 1962: 301. Bowman et al. 1963: 144, fig. 3i. Dunbar 1963: 2 (key), 3, fig. 3. Brusca 1970: 179 –181, fig. 1. Dick 1970: 58, fig. 6. Shih et al. 1971: 65 –66, 165, 221. Yoo 1971: 42 (list), 56. Evans & Sheader 1972: 275 –276. Sanger 1973: 5, 7, 8 & 11 (tables), 10, fig. 3. Sanger 1974: 3 & 5 (tables), 6. Flores & Brusca 1975: 10 –14, figs. 1 & 3. Lorz & Pearcy 1975: 1443 (text fig.), 1444 (table), 1445. Westerhagen 1976: 43–50, fig. 1. Westerhagen & Rosenthal 1976: 669 –673. Harbison et al. 1977: 482 (table). Shih & Laubitz 1978: 50. Laval 1980: passim. Brusca 1981: 10 (list), 41, fig. 7f & 7h. Vinogradov et al. 1982: 282 (key), 283–284, fig. 142. Cahoon et al. 1986: 95 –96. Vinogradov 1992: 325. De Broyer & Jażdżewski 1993: 114 –115. Shih & Chen 1995: 84 (key), 84–86, fig. 50. Spicer & Morritt 1995: 997 –998. Lin et al. 1996: 229 (table). Lavaniegos & Ohman 1999: 495 (table). Vinogradov 1999: 1146 (table), 1185 (key), 1185–1186, fig. 4.108. Lowry 2000: 325 (list). Escobar-Briones et al. 2002: 367 (list). Weslawski & Legezynska 2002: 255 & 257 (tables), 257–258, 260. Brusca & Hendrickx 2005: 150 (list). Gasca et al. 2006: 239. Browne et al. 2007: 819 (table), fig. 4 (phylogenetic tree). Gasca 2009a: 88 (table). Lavaniegos & Hereu 2009: 141, 143, 145, 146 (table), 148, 151 (appendix). Mori et al. 2010: 9 (list). Valencia & Giraldo 2012: 1492 (table). Valencia et al. 2013: 51 (table), 59. Hurt et al. 2013: 31 (table), figs. 1–2 (phylogenetic trees).
non [mis-identification = Hyperoche luetkenides View in CoL ]— Barnard 1932: 276. Hardy & Gunther 1935: 195, 197. Hurley 1955: 144–147, figs. 96–114. Hurley 1960: 112. Hurley 1969: 33, pl. 19 (maps). Lipskaya 1980: 13–14. Ramirez & Vinas 1985: 32–33, figs. 3–4. Jażdżewski & Presler 1988: 63 & 69 (tables), 66, figs. 1–2.). Torres et al. 1994: 210–211 (incl. table).
non [mis-identification =? Hyperoche mediterranea View in CoL ]— Siegfried 1963: 8. Sorarrain et al. 2001: 407–409.
non [mis-identification = Hyperoche capucinus View in CoL ]— Dinofrio 1977: 7 (key), 9, 12; pl. 1, fig. 6; pl. 2, fig. 3.
non [mis-identification = Hyperoche shihi View in CoL ]— Gasca & Haddock 2004: 532 (table), 533, fig. 2B.
Hyperia tauriformis Bate & Westwood, 1868: 519 View in CoL , text fig.
Hyperoche tauriformis View in CoL — Bovallius 1889: 85, 115, text fig. Norman 1900: 130. Norman & Scott 1906: 54 –55. Tattersall 1906: 23. Norman & Brady 1909: 301 (52). Tesch 1911: 180. Tattersall 1913: 21. Bigalow 1926: 165–166. Chevreux 1935: 191.
Hyperia prehensilis Bate & Westwood, 1868: 520 View in CoL , text fig.
Hyperoche prehensilis View in CoL — Bovallius 1887a: 19. Bovallius 1889: 86 (key), 93–94, text fig. Norman 1900: 130 –131.
Metoecus abyssorum Boeck, 1871: 86 –87.
Tauria abyssorum — Boeck 1872: pl. 1, fig. 2. Boeck 1876: 83–84.
Hyperia abyssorum — Bovallius 1885: 17.
Hyperoche abyssorum — Bovallius 1887a: 19. Bovallius 1887b: 564. Bovallius 1889: 86 (key), 92, 94–97, text fig.
Hyperia kroeyeri Bovallius, 1885: 17 .
Hyperoche Kroeyeri View in CoL — Bovallius 1887a: 18. Bovallius 1887b: 564. Bovallius 1889: 86 (key), 87–92, text figs.
Hyperoche kroeyeri View in CoL — Shoemaker 1920: 24 –25.
Hyperoche Kröyeri View in CoL — Sars 1895: 9, pl. 4. Linko 1907: 197. Chevreux & Fage 1925: 405 –406, fig. 404. Stephensen 1913a: 83. Stephensen 1925: 229 (table).
Hyperoche kroyeri — Sheader 1973: 247. Harbison et al. 1977: 482 (table). Laval 1980: 18 (table).
Hyperoche Luetkeni Bovallius 1887a: 19 View in CoL .— Bovallius 1887b: 565, pl. 44, figs. 55–71. Bovallius 1889: 86 (key), 92, 97–105, pl. 7, figs. 1–26.
Hyperoche Lütkeni View in CoL — Vosseler 1901: 55 –56.
Hyperoche luetkeni — Steuer 1911: 674 (key).
Hyperoche leutkeni — Thorsteinson 1941: 88 –90; pl. 8, figs. 83–92.
Hyperoche lutkeni — Laval 1980: 18 (table).
Hyperoche cryptodactylus View in CoL [mis-identification]— Gasca 2009b: 217 (table). Lavaneigos & Hereu 2009: 142, 151 (appendix).
Type material. It seems doubtful that type material of Metoecus medusarum was designated because Kröyer (1838) believed that his material represented the species described by Fabricius (1780). However, Bovallius (1889) seems to have seen some specimens of Kröyer in the ZMUC and determined that they belonged to Hyperoche , but no such specimens could be found in the ZMUC or NRS. The type locality is the west coast of Greenland.
Type material of synonyms. Type material of Hyperia tauriformis and H. prehensilis seems to be lost because no specimens bearing this name could be found in the NHM, London. The type locality for both species is “at Banff, by Mr. Edward”.
Type material of Metoecus abyssorum could not be found in any major European institution and is considered lost. The type locality is Hardangarfjord, Norway.
There is no type material for Hyperia kroeyeri because Bovallius (1885) was only providing a new name for Metoecus medusarum Kröyer, 1838 (thought to belong to Hyperia ) because Hyperia medusarum ( Müller, 1776) has precedence but is a different species.
Some syntypes of Hyperoche luetkeni are in the ZMUC. One female ( CRU- 7071), from the south-eastern tip of Greenland [57°49’N 35°24’W], “Bang 1868”, and 14 females and 4 males ( CRU- 9243), from Jakobshavn, West Greenland, “Borch 1859”. There are also some unregistered specimens in the NRS which may represent type material, and it is likely that Bovallius (1887b) used some of it to illustrate his species. All the material is on microscope slides. One lot (two slides) from “Spitzbergen, 1881” and the remainder from off Greenland as follows. One slide [58°N 50°W], one slide [67°35’N] and six slides from the same locality as ZMUC CRU- 7071.
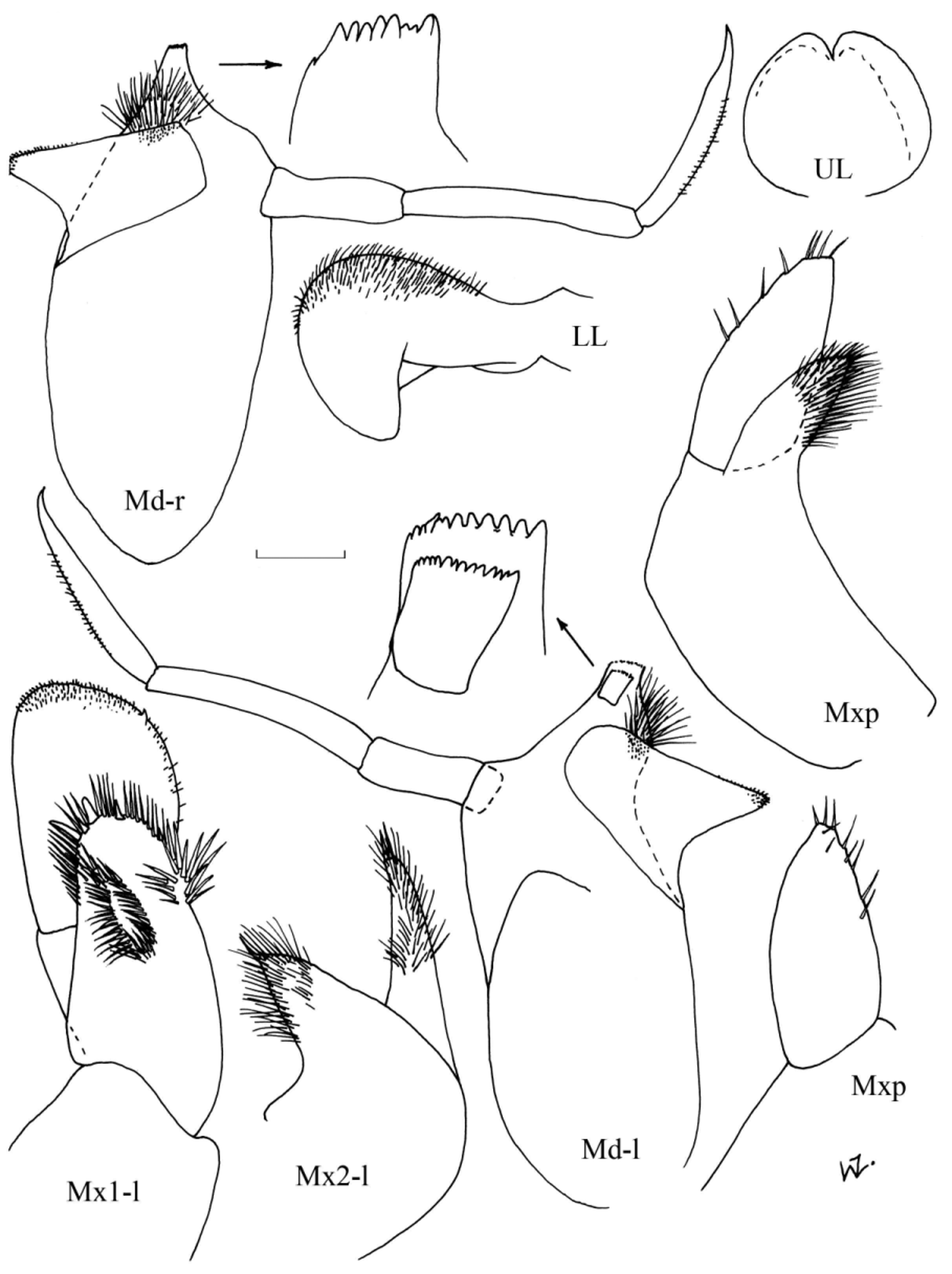
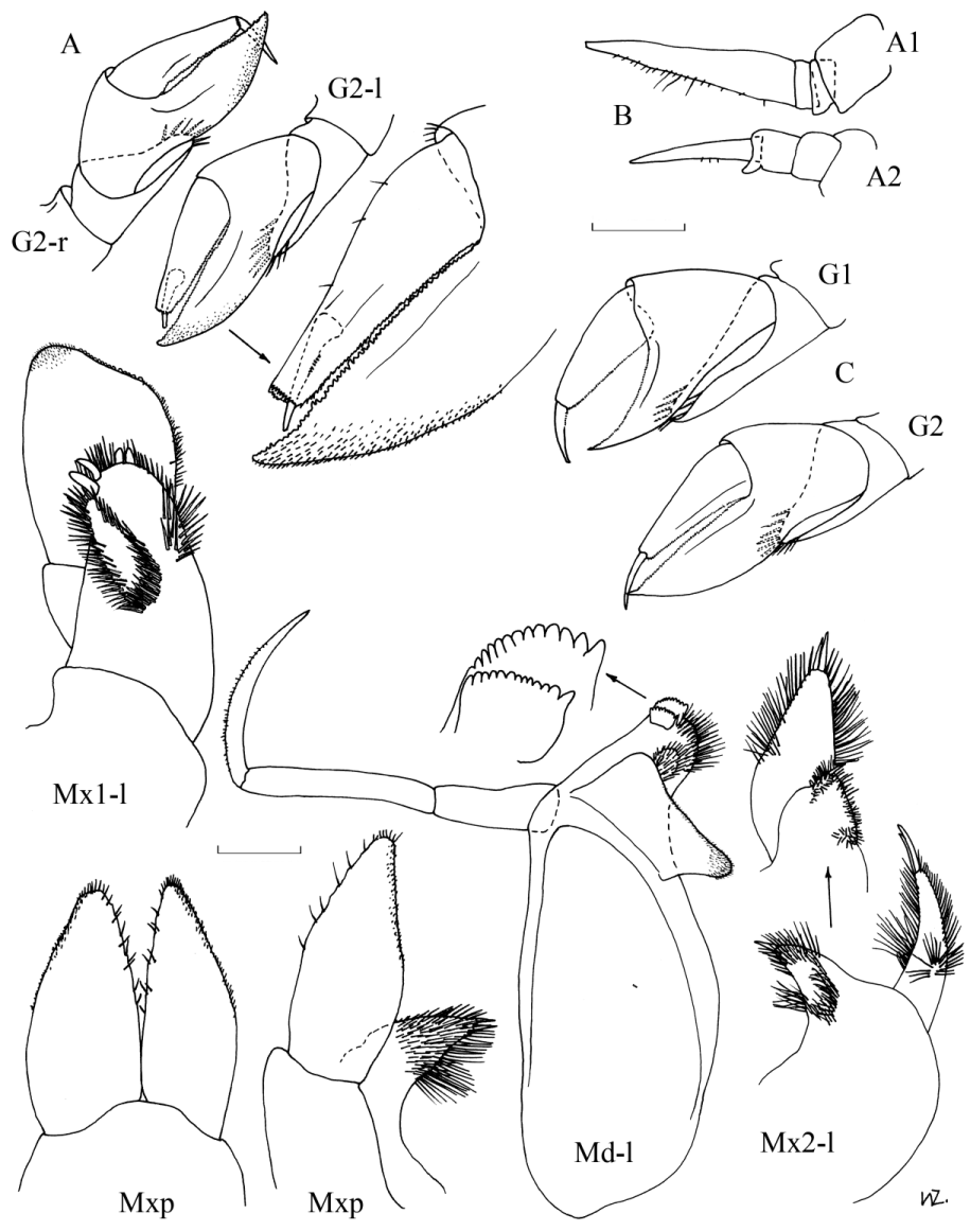
Diagnosis. Females: Sexually mature at about 7.0 mm (reports of some Arctic specimens reaching 15.0 mm remains to be confirmed). Antennae 1 as long as head, about 1.6 x A2. Head length equal to first two pereonites combined. Pereon globular, length about 1.5 x pleon. Gnathopod 1; basis marginally shorter than remaining articles combined, relatively broad with maximum width about 0.6 x length; merus spoon-shaped, projecting under carpus, almost to base of propodus, with fringe of setae on distal margin; carpal process extends just past distal margin of propodus, anterior margin denticulate; posterior and distal margin of propodus also denticulate; dactylus slightly curved, posterior margin finely denticulate, length almost 0.4 x propodus. Gnathopod 2 slightly longer and more slender than G1 but similar in structure except for slightly shorter spoon-shaped process of merus. Pereopods 3 & 4 similar in length to P5 & 6, or marginally shorter. Pereopod 3; basis length 2.5 x merus; carpus with postero-distal corner produced into slight tooth with denticulate margins, length 1.2 x merus and 0.8 x propodus; posterior margin of propodus denticulate; dactylus length slightly more than 0.2 x propodus. Pereopod 4 slightly more slender than P3 but similar in structure except the basis is relatively shorter, the carpus and propodus are relatively longer and the postero-distal corner of the merus is rounded. Pereopods 5 & 6 are similar in size and structure; basis length about twice merus; carpus length about 1.3 x merus, slightly shorter than propodus; dactylus length 0.3 x propodus. Pereopod 7; coxa fused with pereonite; similar in structure to P6 but slightly shorter because merus is relatively shorter. Epimeral plates with postero-distal corner produced into small point. Uropod 1; peduncle reaching to limit of peduncle of U2 and to about half peduncle of U3; inner ramus marginally longer than outer, slightly shorter than peduncle. Uropod 2; inner ramus slightly shorter than peduncle, about 1.5 x length of outer ramus. Uropod 3; inner ramus marginally longer and wider than outer, about half-length peduncle. Telson triangular, marginally longer than wide, slightly shorter than half of peduncle of U3.
Colour in life: according to Shoemaker (1920), based on specimens from the Alaskan Arctic. “Central areas of eyes duck green. Dorsal parts of the body segments apricot orange. Sides of body, sideplates, gnathopods, peraeopods, pleopods, uropods and telson splotched with apricot orange. Rest of animal translucent.”
Males: Sexually mature at about 7–8 mm. Antennae as long as entire animal. Pereon and pleon slender, of similar length. Appendages generally more slender than in females, especially the gnathopoda, otherwise very similar in structure and relative lengths of articles, except for the following minor variations. Gnathopoda with merus not projected as far under the carpus. Pereopods 3 & 4 with postero-distal corner of the carpus less prominent. Epimeral plates relatively much longer and deeper. Uropod 1; peduncle extends to just past limit of peduncle of U2, to about 0.6 x peduncle of U3; both rami with characteristic proximal excavation. Telson as wide as long, about 0.4 x length of peduncle of U3.
Material examined. Some syntypes of Hyperoche luetkeni as detailed above and the following additional material.
N.W. Atlantic: Iceland / Greenland region, including Davis Strait; expeditions of Godthaab, Thor, Ingolf, Dana and others; 134 females, 85 males (82 lots; ZMUC), all near surface. N.E. Atlantic: Vicinity of Denmark and the North Sea; 160 females, 54 males, 2 juveniles (34 lots; ZMUC), all near surface. Four lots ( ZMUC), near the Faeroe Islands; 4 females [62°10’N 07°30’W], Vestmanna, surface, 11 July 1900; two females [62°25’N 07°40’W], Dana stn. 2228, 65 mw, 1 August 1924; female [61°32.5’N 06°47’W], Dana stn. 2241, 40 mw, 3 August 1924; two females [61°55’N 07°30’W], Dana stn. 7661–1, 118– 120 m, 14 June 1951. Near the English Channel ( ZMUC); female [49°17’N 04°13’W], Thor stn. 1, 25 mw, 28 November 1908; 3 females [47°01’N 05°48’W], Thor stn. 76, 25 & 65 mw, 10 March 1909. 3 males ( ZMUC), mid-Atlantic [48°43’N 40°45’W], Thor stn. 247, surface, 19 September 1910. N.E. Pacific: Fifteen females, 9 males (11 lots; SAMA C7913–7923), vicinity of Vancouver Island, British Columbia [range about, 48°–52°N 124°31’– 129°42’W], Line P and La Perouse Monitoring Program, 0–250 m, ex. M. Galbraith. Male ( SAMA C7924), off British Columbia [49°58.10’N 144°56.20’W], Cruise 9824, stn. 98090, 1200 m, 8 July 1998. Male ( SAMA C7925], off British Columbia [50°00.32’N 144°54.07’W], Cruise 0 229, stn. OSO 250, 1200 m, 18 July 2002. Six males ( SAMA C7926), stomach of Pink Salmon [51°N 145°W], Cruise 2012, stn. OSO 134, 10 July 2000. Female, 2 males ( SAMA C7927), south of Alaskan Peninsula [50°00.50’N 165°00.20’W], 150 m, September 1999. Female ( SAMA C7928), north-west of Graham Island [54°20.60’N 133°17.80’W], Cruise 9816 PBS, stn. 2200, 16 June 1998. Male ( SAMA C7929), off Vancouver Is. [50°N 145°W], Cruise IOS 0 311 stn. P 26, 250 m, 3 June 2003. Female, male ( SAMA C7930), off San Francisco [37°53’N 123°25’W], NORPAC, 50–0 m, September 2005. Female ( SAMA C7931), Californian Transition Zone, 76– 51 m, Cruise CTZ 88, stn. 30, 10 June 1988.
Remarks. This is a medium-sized species, reaching sexual maturity at about 7–8 mm, although Vinogradov et al. (1982) note that some Arctic specimens reach 15 mm, but this could not be confirmed from the literature or from specimens examined.
This species is very difficult to distinguish from H. luetkenides . According to Walker (1906), who based his description on a unique male, the main character distinguishing H. luetkenides from H. medusarum is the ornamentation of the distal margin of the merus and carpus of pereopods 3 & 4, which in H. luetkenides is not denticulate, just armed with setae. However, in males of Hyperoche the articles of the pereopoda are usually less ornamented than in females, and in males of H. medusarum the distal margin of the merus and carpus of pereopods 3 & 4 is very finely serrated, giving the appearance of setae. Thus, this character cannot be used to separate these two species. Hence there are a number of Southern Hemisphere records of H. medusarum most of which are here considered to represent H. luetkenides . Compared to H. medusarum , females of H. luetkenides generally have the gnathopoda, pereopoda and uropoda with slightly more robust articles; the spoon-shaped merus and carpal process of the gnathopoda extends slightly further; the peduncle of U1 does not reach the limit of the peduncle of U2 and the second antennae seem to be relatively shorter, about 1.3 x A2 (about 1.6 x for H. medusarum ). Hyperoche luetkenides is also a much larger species, sexually mature at about 14–18 mm with a maximum size of about 20 mm.
During this study a few male specimens were found to have some pereopoda with the dactyls retracted, sometimes only partially ( Fig. 3 View FIGURE 3 ), and usually only on one side. This character probably also occurs in females because it has been found in females of H. luetkenides , occurring in one of the second gnathopoda ( Fig. 9 View FIGURE 9 ). This character does not seem to be common but probably occurs sporadically in all species of Hyperoche . It seems unlikely that this character is an artefact of preservation because it has not been noted in any other hyperiid that normally has non-retractile dactyls.
This species has been recorded as an associate of the following medusa; Chrysaora sp. and other large medusa ( Gosse 1853 and many subsequent authors in general), Aurelia aurita ( Stephensen 1923a) , Cyanea capillata & Tima formosa ( Bowman et al. 1963) , and Mitrocoma cellularia ( Gasca et al. 2006) . It has also been found on the ctenophores Beroe forskalii ( Stephensen 1923a, Pirlot 1939), Beroe cucumis & Pleurobrachia pileus ( Sheader 1973, Evans & Sheader 1972), Pleurobrachia bachei ( Brusca 1970, Flores & Brusca 1975), Beroe ovata & Mnemiopsis leidyi ( Cahoon et al. 1986) and Mnemiopsis mccradyi ( Sorarrain et al. 2001) . In addition Westernhagen (1976) records it from hydromedusae of the genera Sarsia, Tiaropsis and Phialidium or Polyorchis .
Distribution. This species is relatively abundant in the colder parts of the Northern Hemisphere; found mainly in near-surface waters. In the North Atlantic it mainly inhabits Arctic waters, rarely venturing further south than about 50°N. In the North Pacific it is also more common in Arctic waters but ranges further south to about 30°N; in the west to the East China Sea, and sometimes beyond, and in the east to the Gulf of California.
It has not been recorded from the Indian Ocean, and it is unlikely to occur there because it seems to be a predominantly Arctic species.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hyperoche medusarum ( Kröyer, 1838 )
| Zeidler, Wolfgang 2015 |
Hyperoche cryptodactylus
| Gasca 2009: 217 |
| Hereu 2009: 142 |
Hyperoche lutkeni
| Laval 1980: 18 |
Hyperoche kroyeri
| Laval 1980: 18 |
| Harbison 1977: 482 |
| Sheader 1973: 247 |
Hyperoche leutkeni
| Thorsteinson 1941: 88 |
Hyperoche kroeyeri
| Shoemaker 1920: 24 |
Hyperoche luetkeni
| Steuer 1911: 674 |
Hyperoche Lütkeni
| Vosseler 1901: 55 |
Hyperoche Kröyeri
| Chevreux 1925: 405 |
| Stephensen 1925: 229 |
| Stephensen 1913: 83 |
| Linko 1907: 197 |
| Sars 1895: 9 |
Hyperoche tauriformis
| Chevreux 1935: 191 |
| Tattersall 1913: 21 |
| Tesch 1911: 180 |
| Norman 1909: 301 |
| Norman 1906: 54 |
| Tattersall 1906: 23 |
| Norman 1900: 130 |
| Bovallius 1889: 85 |
Hyperoche medusarum
| Valencia 2013: 51 |
| Hurt 2013: 31 |
| Valencia 2012: 1492 |
| Mori 2010: 9 |
| Gasca 2009: 88 |
| Lavaniegos 2009: 141 |
| Browne 2007: 819 |
| Gasca 2006: 239 |
| Brusca 2005: 150 |
| Escobar-Briones 2002: 367 |
| Weslawski 2002: 255 |
| Lowry 2000: 325 |
| Lavaniegos 1999: 495 |
| Vinogradov 1999: 1146 |
| Lin 1996: 229 |
| Shih 1995: 84 |
| Spicer 1995: 997 |
| De 1993: 114 |
| Cahoon 1986: 95 |
| Vinogradov 1982: 282 |
| Brusca 1981: 10 |
| Shih 1978: 50 |
| Harbison 1977: 482 |
| Westerhagen 1976: 669 |
| Flores 1975: 10 |
| Lorz 1975: 1443 |
| Sanger 1974: 3 |
| Sanger 1973: 5 |
| Evans 1972: 275 |
| Shih 1971: 65 |
| Yoo 1971: 42 |
| Brusca 1970: 179 |
| Dick 1970: 58 |
| Bowman 1963: 144 |
| Dunbar 1963: 2 |
| Grice 1962: 301 |
| Irie 1958: 107 |
| Irie 1957: 10 |
| Hurley 1956: 14 |
| Vinogradov 1956: 211 |
| Bulycheva 1955: 1043 |
| Dunbar 1954: 782 |
| Bousfield 1951: 139 |
| Stephensen 1944: 9 |
| Stephensen 1942: 458 |
| Stephensen 1940: 5 |
| Behning 1939: 361 |
| Pirlot 1939: 38 |
| Stephensen 1933: 61 |
| Stephensen 1932: 375 |
| Barnard 1930: 415 |
| Pirlot 1929: 118 |
| Stephensen 1929: 42 |
| Stephensen 1928: 30 |
| Schellenberg 1927: 630 |
| Stephensen 1924: 79 |
| Stephensen 1923: 13 |
| Stephensen 1923: 7 |
| Stephensen 1913: 99 |
| Stephensen 1912: 557 |
| Stappers 1911: 77 |
| Hansen 1888: 58 |
| Stebbing 1888: 1399 |
Hyperoche prehensilis
| Norman 1900: 130 |
| Bovallius 1889: 86 |
| Bovallius 1887: 19 |
Hyperoche abyssorum
| Bovallius 1889: 86 |
| Bovallius 1887: 19 |
| Bovallius 1887: 564 |
Hyperoche
| Bovallius 1889: 86 |
| Bovallius 1887: 18 |
| Bovallius 1887: 564 |
Hyperoche Luetkeni Bovallius 1887a : 19
| Bovallius 1889: 86 |
| Bovallius 1887: 19 |
| Bovallius 1887: 565 |
Hyperia abyssorum
| Bovallius 1885: 17 |
Hyperia kroeyeri
| Bovallius 1885: 17 |
Tauria medusarum
| Sars 1882: 75 |
Metoecus abyssorum
| Boeck 1871: 86 |
Hyperia tauriformis
| Bate 1868: 519 |
Hyperia prehensilis
| Bate 1868: 520 |
Metoicus medusarum
| Kinahan 1859: 83 |
Metoecus medusarum Kröyer, 1838 : 288
| Boeck 1871: 86 |
| White 1857: 207 |
| Gosse 1853: 367 |
| White 1847: 91 |
| Milne-Edwards 1840: 78 |
| Kroyer 1838: 288 |