
Himalotrechus, Belousov & Kabak & Schmidt, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4544.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:03B32349-24F9-465B-AA7E-0B4DCA339B96 |
|
DOI |
https://doi.org/10.5281/zenodo.5942608 |
|
persistent identifier |
https://treatment.plazi.org/id/AE5D87F2-2D2B-FFB3-FF15-90F4FC60FC5B |
|
treatment provided by |
Plazi |
|
scientific name |
Himalotrechus |
| status |
gen. nov. |
Himalotrechus View in CoL gen. n.
Type species: Himalotrechus humeratus sp. n.
Description. Rather large-sized, body length of the only known species is more than 4.5 mm. Body extraordinarily convex and broadly ovate, elytra suborbicular, more narrowed apically than basally, widest near humeral level; pronotum markedly wider at base than at apex; head trapezoid, narrowed anteriad; legs and antennae rather short ( Fig. 1).
Color: dark amber-brownish.
Microsculpture: body surface completely covered with sculpticells visible at magnification of 56x.
Head: trapezoid, rather narrow in anterior portion, frontal furrows continuous. Eyes small, subconvex, much shorter than the glabrous tempora. Two supraorbital setiferous pores. Labrum sexsetose, its anterior margin concave. Mandibles slender, tooth of right mandible with distinct premolar and anterior denticle of retinaculum displaced distally (tridentate type) ( Fig. 2 View FIGURES 2–3 ). Labial tooth moderately salient, rounded or truncate. Six submental setae. Mentum and submentum fused, without distinct suture between them. Maxillary palpi elongate, glabrous ( Fig. 3 View FIGURES 2–3 ). Penultimate segment of labial palpi bisetose. Glossa markedly salient medially, with two median setae much longer than the lateral ones. Paraglossae moderately long, slender, slightly curved, distinctly pubescent.
Pronotum: large, slightly transverse trapezoid, basal margin significantly longer than anterior one. Hind angles subrectangular to obtuse angular, pointed apically. Lateral margins narrowly reflexed anteriorly, very markedly reflexed posteriorly, lateral groove narrow anteriorly, widened posteriorly. Prebasal transverse impression shallow. Two lateral setiferous pores on each side of pronotum, the posterior one just in the hind angles.
Elytra: very convex, broadly ovate, with maximum width in anterior third. Prehumeral margin almost perpendicular to elytral suture, with a tuberculum near the mesothoracic constriction. Humeri prominent but rounded. Posterior portion of elytra narrowed towards apex. Lateral groove rather wide; margins moderately reflexed, more markedly in the humeral area; lateral border disappearing anteriorly at level of stria 5. All elytral striae well-impressed and very coarsely punctured though becoming very shallow at base and on apical slope. Parascutellar pore present, parascutellar striole developed. Apical striole well-impressed, directed to stria 5 anteriorly. Two discal pores in anterior half of elytra, both attached to stria 3; preapical pore lacking. Two apical pores of which angulo-apical one is much smaller. Umbilicate series well-aggregated, only pore 5 slightly shifted from lateral margin.
Ventral side: metepisternites smooth, approximately 1.5 times as long as wide. Only one paramedian seta on each side of the abdominal sternites. Anal sternite with two setae in male and four setae in female.
Legs: Protibia markedly grooved on exterior surface, without pubescence on anterior surface. Male protarsi with two basal segments dilated. Profemora simple in males, without ventral projection and additional setae.
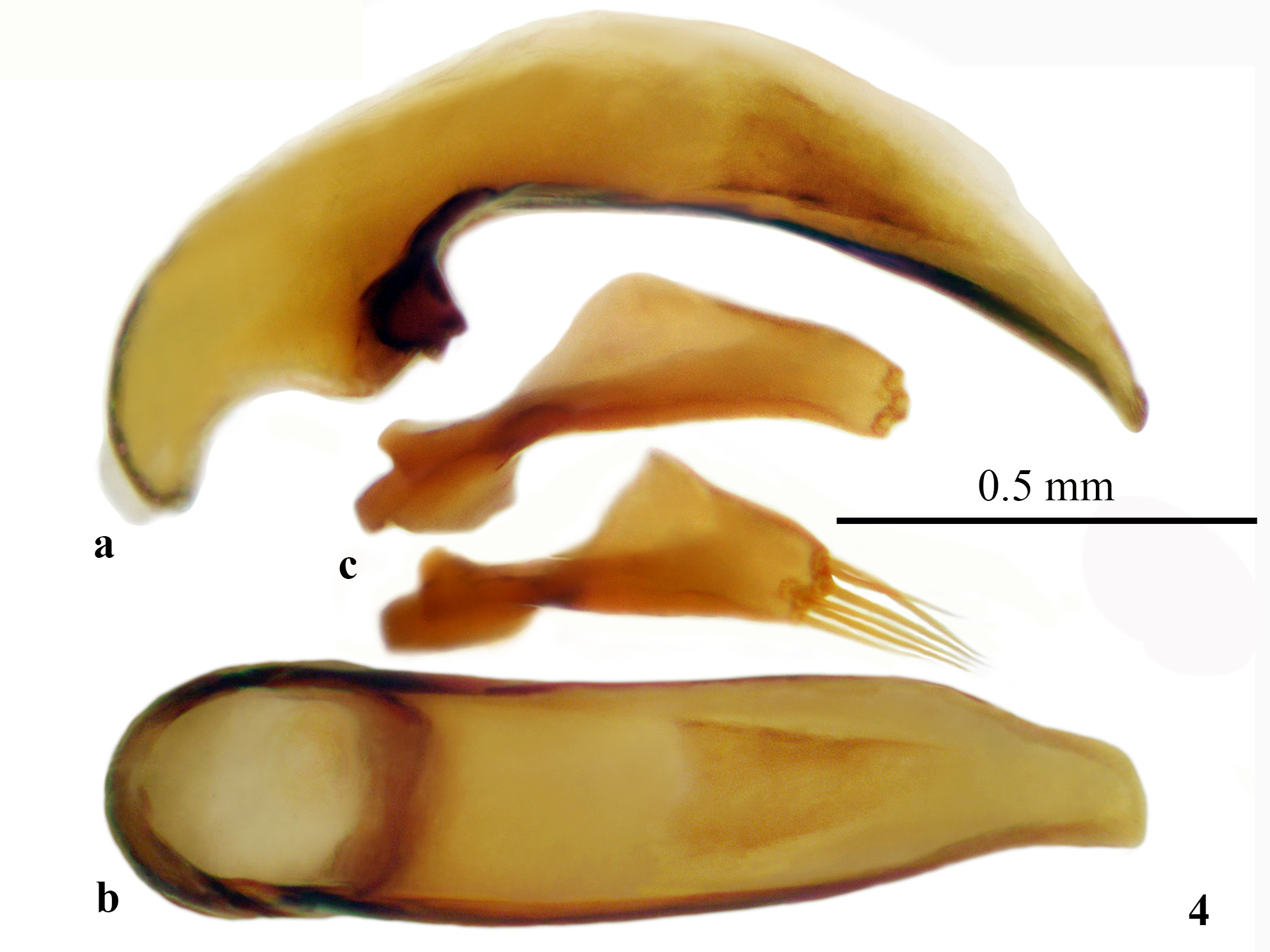
Aedeagus: median lobe of moderate size, rather thick, with apex slightly attenuated ventrally ( Fig. 4 View FIGURE 4 ). In dorsal view, median lobe subparallel-sided for most of its length, without dilation, only the apical lamella narrowed. Sagittal aileron small but distinct. Endophallus armature poorly sclerotized, consisting of asymmetrical scaly areas confined to the apical half of the median lobe. Parameres robust, each bearing five to six apical setae.
Comparative notes. Externally the new genus resembles representatives of the genera Agonotrechus and Bhutanotrechus of the Agonotrechus -complex sensu Uéno (1977), which are also known to occur in the Eastern Himalaya. All these taxa share the following characters: body broadly ovate, head elongate and narrow, eyes rather small, approximately as long as tempora (at least, in wingless species), pronotum large, with lateral margins markedly expanded and reflexed posteriorly and elytra very wide and convex. Despite external similarities, the new genus does not appear to be directly related to any lineage of the Agonotrechus -complex, as demonstrated by the lack of pubescence on the anterior surface of the distal part of the protibia. This character is of great importance for taxonomy of the tribe Trechini and, it is the only one that distinguishes Trechus and Agonotrechus related taxa in many cases (e.g., see discussion of the systematic position of Trechus gitzeni in Belousov & Kabak, 2001 ).
Actually Himalotrechus View in CoL gen. n. appeared to be related to some taxa of the Homaloderini evolutionary lineage (sensu Jeannel, 1927). The latter, though not recognized by most modern carabidologists as a distinct tribe or subtribe, embraces a few distinct lineages, of which, at least, some should be considered as natural groups. In the Oriental Region, the following lineages are worth noting in relation to the genus under discussion: Sumatrechus Deuve, 2005 View in CoL from Sumatra and Luzonotrechus Uéno, 1979 View in CoL from the Philippines ( Deuve 2005; Uéno 1979).
At the present state of knowledge, Himalotrechus View in CoL gen. n. seems to be a representative of one of the Homaloderini evolutionary lines in the sense of Uéno (1979) based on the following characters: i) elytra broadly ovate (much wider than pronotum); ii) head rather narrow compared to pronotum; iii) mandibles slender, slightly curved; iv) tooth on the right mandible with a well-defined premolar separated from distal denticle of retinaculum by a long and deep incision. However, the new genus readily differs from the genus Sumatrechus View in CoL in the following characters: pronotum with posterior lateral setiferous pore; elytra with two discal setiferous pores, and without preapical pore; lateral margins of pronotum markedly expanded and reflexed in posterior part; endophallus armature poorly sclerotized, without well-defined scaly areas.
The differences between the new genus and known taxa of Luzonotrechus are less obvious. The original diagnosis of this genus ( Uéno 1979) was substantially modified later to incorporate newly described species ( Uéno 1987). This latter diagnosis shows a high degree of morphological variation in members of Luzonotrechus quite unusual for most Trechini genera: the male protarsi are simple in some species, with one or two basal segments dilated in others; laterobasal seta of pronotum lacking or present; elytral striation complete or reduced laterally; stria 2 either reaching elytral apex without anastomosis with stria 3 or with apical cross (striae 2 and 3 joining each other on apical slope) as in most Trechini ; one or two discal setiferous pores present; profemora either simple or sexually dimorphic (angulate or denticulate ventrally and bearing one or two hairs on the angulation in males), etc. Therefore, of all essential diagnostic characters used initially, only the three following features are kept as valid: the very characteristic shape of the male genitalia (median lobe very slender at base, sagittal aileron large, median lobe inflated medially or in preapical portion), preapical pore absent, and premolar well-defined on the right mandible. The latter character is assumed to be plesiomorphic and is shared by all lineages of Homaloderini sensu Uéno (1979). Himalotrechus gen. n. shares with Luzonotrechus the presence of the premolar and absence of the preapical pore but differs in the male genitalia structure which disagrees with those of Luzonotrechus in all essential points. Uéno (1987) considered the male genital structures as the most essential character for recognizing the true systematic position of “ Luzonotrechus ” orousseti Perrault, 1982: “the species has the ordinarily shaped male genitalia… this species seemed to me to belong to the group of Trechus bakeri ” ( Uéno 1987: p. 123–124). Additionally, it is worthy to note that the combination of characters observed in Himalotrechus gen. n. is not found in any species of the genus Luzonotrechus .
The problem of Trechini bidentati and Trechini tridentati was recently raised again by T. Deuve, D. Kavanaugh and H. Liang (2016). Studying extensive trechine material collected during multi-year international projects in southern Yunnan, they concluded that the mandibular dentition is a character of primary importance for understanding the local trechine fauna. Based mainly on this character, they described 3 new genera with many new species and some known species moved to these genera from the genus Trechus Clairville, 1806 . Apart from very useful discussion of evolutionary trends in the structure of mandibular dentition, they proposed to group genera with distinct premolar and retinaculum characterized by anterior denticle displaced distally into the Epaphiopsis Complex. Such an approach seems to be opportune since it allows avoiding the term Homaloderini while preserving the focus on this important character. Our data are in line with these views.
According to our opinion, Himalotrechus gen. n. is most closely related to just these three genera. Especially, Trechepaphiama Deuve & Kavanaugh, 2016 with only one known species— T. gaoligong Deuve & Kavanaugh, 2016 —seems to be very similar to Himalotrechus gen. n. Both genera share the same body shape: elytra convex, suborbicular, more narrowed apically than basally, with prominent humeri; head trapezoid, narrowed anteriad; pronotum wider at base that at apex, with sides barely sinuate before hind angles; very similar pattern of the mandibular dentition (the distal denticle of retinaculum approximately in the middle between the premolar and mandibular tip) and glabrous tempora. However, the elytral chaetotaxy shows significant differences: discal setiferous pores lack, preapical pore present in Trechepaphiama while two discal pores present and the preapical pore missing in Himalotrechus gen. n. Thus, the two genera under consideration demonstrate opposite patterns of the elytral chaetotaxy and their direct relationships may be questioned. Apart from the above characters, Himalotrechus gen. n. differs in some less important characters: elytral striae deeply impressed and coarsely punctate, size bigger and color darker.
In contrast to Trechepaphiama, two known species of Epaphiotrechus Deuve & Kavanaugh, 2016 strongly differ from Himalotrechus gen. n. in their habitus: body shape elongate, rather flat; pronotum approximately as wide at base as at apex; elytral striae shallow, finely punctate. The elytral chaetotaxy of this genus is typical for most Trechini : two discal setiferous pores and one preapical pore in the apical cross of striae 2 and 3. Thus, in regard to the elytral chaetotaxy, Himalotrechus gen. n. differs from Epaphiotrechus only in the absence of the preapical pore. The main difference between the two genera under consideration is the shape of the mandibular tooth. Epaphiotrechus shows an intermediate evolutionary state of the bidentate type ( Deuve & al. 2016) while Himalotrechus gen. n. has a typical tridentate type with an isolated premolar and the anterior denticle of retinaculum markedly displaced distally.
Finally, members of the genus Trechepaphiopsis Deuve & Kavanaugh, 2016, apart from their Trechus -like appearance, differs readily from Himalotrechus gen. n. in having tempora slightly pubescent, elytra with only one or no discal seta and elytral striae slightly impressed and finely punctate. This genus seems to be more remotely related to Himalotrechus gen. n. than two other genera considered above.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Trechini |
Himalotrechus
| Belousov, Igor A., Kabak, Ilya I. & Schmidt, Joachim 2019 |
Himalotrechus
| Belousov & Kabak & Schmidt 2019 |
Himalotrechus
| Belousov & Kabak & Schmidt 2019 |
Sumatrechus
| Deuve 2005 |
Sumatrechus
| Deuve 2005 |
Luzonotrechus Uéno, 1979
| Ueno 1979 |