
Heterometrus swammerdami Simon, 1872
|
publication ID |
https://doi.org/ 10.18590/euscorpius.2016.vol2016.iss220.1 |
|
publication LSID |
lsid:zoobank.org:pub:DD0DF45D-F63A-4AA2-8EFF-03CF99E297EF |
|
DOI |
https://doi.org/10.5281/zenodo.7124602 |
|
persistent identifier |
https://treatment.plazi.org/id/B07187DF-A867-FFE2-FECA-F9059AA2FAFF |
|
treatment provided by |
Felipe |
|
scientific name |
Heterometrus swammerdami Simon, 1872 |
| status |
|
Heterometrus swammerdami Simon, 1872
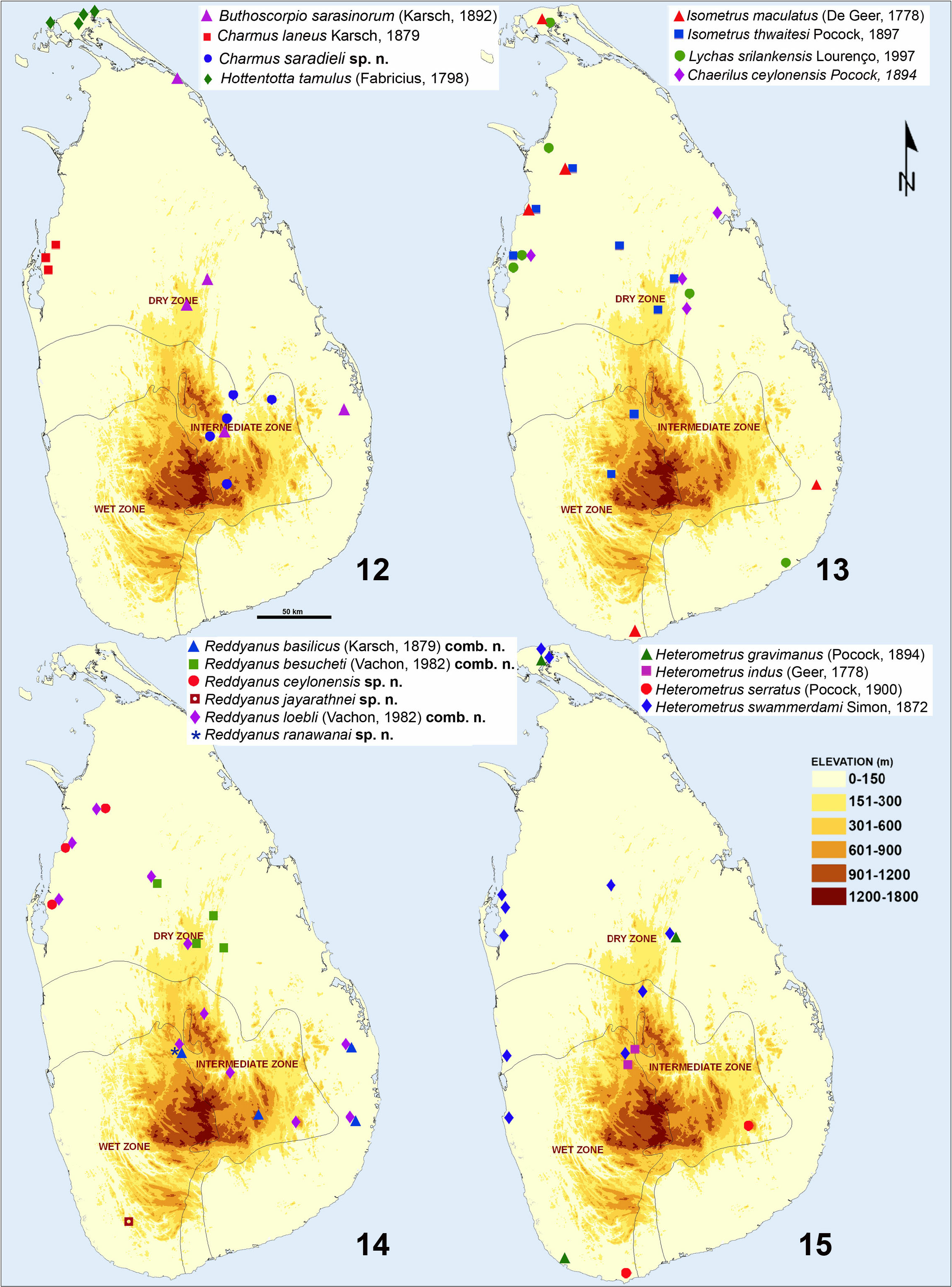
( Figs. 15 View Figures 12–15 , 535–546 View Figures 535–536 View Figures 537–538 View Figures 539–543 View Figures 544–546 )
Heterometrus swammerdami Simon, 1872: 56–59 ; Fet, 2000: 443–444 (complete reference list until 1998); Kovařík, 200 4: 42–44, fig. 28 (complete reference list until 2004); Kovařík, 2009: 44, 93, 113, figs. 139–145, 275–283; Veronika & al., 2013: 72–73, figs. 1, 3–8, tab. 1.
Heterometrus (Gigantometrus) swammerdami: Tikader & Bastawade, 1983: 562–567 , figs. 1497–1510;
= Heterometrus (Gigantometrus) swammerdami titanicus Couzijn, 1981: 165–167 , figs. 4f, 55, 56 (syn. by Kovařík, 2004: 42).
TYPE LOCALITY AND TYPE REPOSITORY. East Indies; MNHN.
SRI LANKAN MATERIAL EXAMINED. Sri Lanka, North Western Province, Puttalam , IV.1994, 1♂ 1♀ 1juv., FKCP, leg. P. Senft ; Central Province, Kandy District, Hantana ( Peradeniya ), University land, 07°14'54.7"N 080°36'54"E, 760 m a.s.l. ( Locality 15CA, Fig. 575 View Figures 575–576 ), 19. –21. IV.2015, 1♀, UPSL, leg. Kovařík et al.; GoogleMaps Central Province, Matale District, Maragamuwa / Kumaragala , 0 7°41'58.5"N 080°42'26.7"E, 662 m a.s.l. ( Locality 15CB, Fig. 576 View Figures 575–576 ), 21. IV.2015, 1♀ ( Figs. 535–543 View Figures 535–536 View Figures 537–538 View Figures 539–543 ), FKCP, leg. Kovařík et al.; GoogleMaps North Central Province, Polonnaruwa District, Giritale, 08°01'26.0"N 080°54' 37.2"E, 233 m a.s.l. ( Locality 15CE, Figs. 579–580 View Figures 579–581 ), 22. –23. IV.2015, 1juv. ( Fig. 545 View Figures 544–546 ), FKCP, 3juvs., UPSL, leg. Kovařík et al.; GoogleMaps Northern Province, Jaffna District, 09°42'51.6"N 080°04'44.8"E, 19 m a.s.l. ( Locality 15CJ, Fig. 587 View Figures 585–587 ), 26. –27. IV.2015, 1♀, FKCP, leg. Kovařík et al.; GoogleMaps North Central Province, Anuradhapura District, Mihintale , 08°20'51.8"N 080°30'27.7"E, 156 m a.s.l. (Locality 15CL, Fig. 589 View Figures 588–589 ), 27. –28. IV.2015, photos only, leg. Kovařík et al. GoogleMaps ; North Central Province, Puttalam District, Eluwankulam , 08° 1 2'35.1"N 079°51'32"E, 52 m a.s.l. (Locality 15CN, Fig. 591 View Figures 590–591 ), 28. IV.2 015, photos only, leg. Kovařík et al.; GoogleMaps North Central Province, Puttalam District, Eluwankulam, 08°17'15"N 079°50' 38.7"E, 38 m a.s.l. ( Locality 15CO, Fig. 592 View Figures 592–594 ), 28. IV.2 015, 1juv., UPSL, leg. Kovařík et al. GoogleMaps
DIAGNOSIS. Total length 1 28–176 mm long. Base color uniformly reddish brown to reddish black. Juveniles may be red with yellow telson. Pectinal teeth number 16–20. Sexual dimorphism in proportions of pedipalps not noticeable. Chela strongly lobiform, length/ width ratio 1.6–1.8 in both sexes. Entire manus covered by large, rounded granulae that do not form true carinae. Pedipalp patella without pronounced internal tubercle. Carapace with disc smooth, margins and posterior portion granulate, anterior portion granulate and tuberculate; occasionally entire surface sparsely granulate. Fifth segment of metasoma longer than pedipalp femur, fourth segment of metasoma approximately as long as pedipalp femur. Telson bulbous, vesicle as long as or longer than aculeus.
NOTE. For photos of the male see figs. 139–142 and 275 in Kovařík, 2009: 93 and 113.
DISTRIBUTION. India and Sri Lanka.










| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Heterometrus swammerdami Simon, 1872
| Kovařík, František, Lowe, Graeme, Ranawana, Kithsiri B., Hoferek, David & Š, V. A. 2016 |
Heterometrus (Gigantometrus) swammerdami titanicus
| KOVARIK 2004: 42 |
Heterometrus (Gigantometrus) swammerdami: Tikader & Bastawade, 1983: 562–567
| TIKADER B. K. & D. B. BASTAWADE 1983: 567 |
Heterometrus swammerdami
| VERONIKA, K. & K. AKILAN & A. MURUGAN- ANTHAN & T. ESWARAMOHAN 2013: 72 |
| KOVARIK, F. 2009: 44 |
| Fet 2000: 443 - 444 |
| SIMON, E. 1872: 59 |