
Lochmanolenellus subquadratus, Webster, Mark & Bohach, Lisa L., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3824.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:023D78D0-4182-48D2-BAEB-CDA6473CF585 |
|
DOI |
https://doi.org/10.5281/zenodo.6129732 |
|
persistent identifier |
https://treatment.plazi.org/id/B10C8793-FFD7-FFBB-61B5-F901FA688776 |
|
treatment provided by |
Plazi |
|
scientific name |
Lochmanolenellus subquadratus |
| status |
sp. nov. |
Lochmanolenellus subquadratus n. sp.
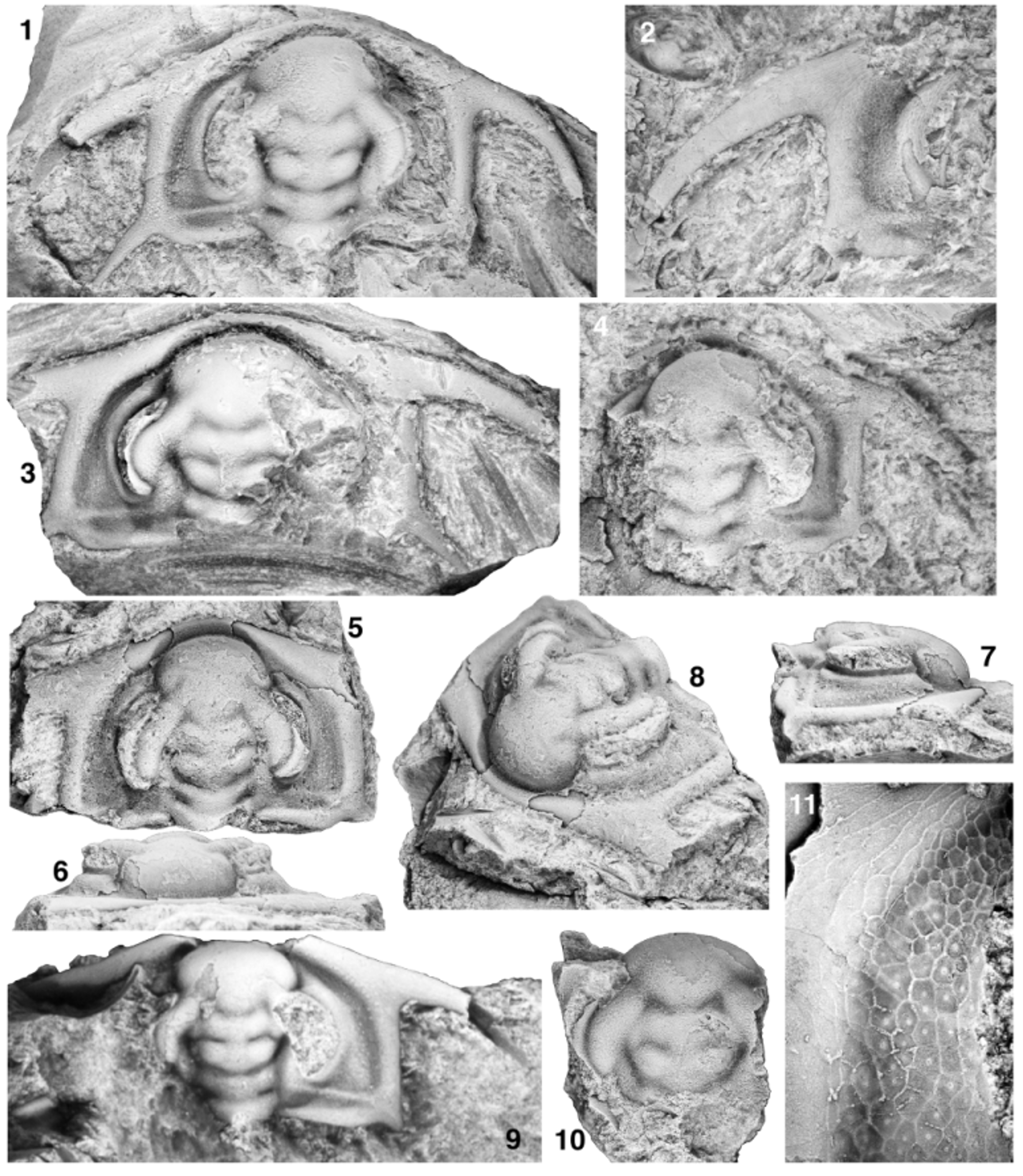
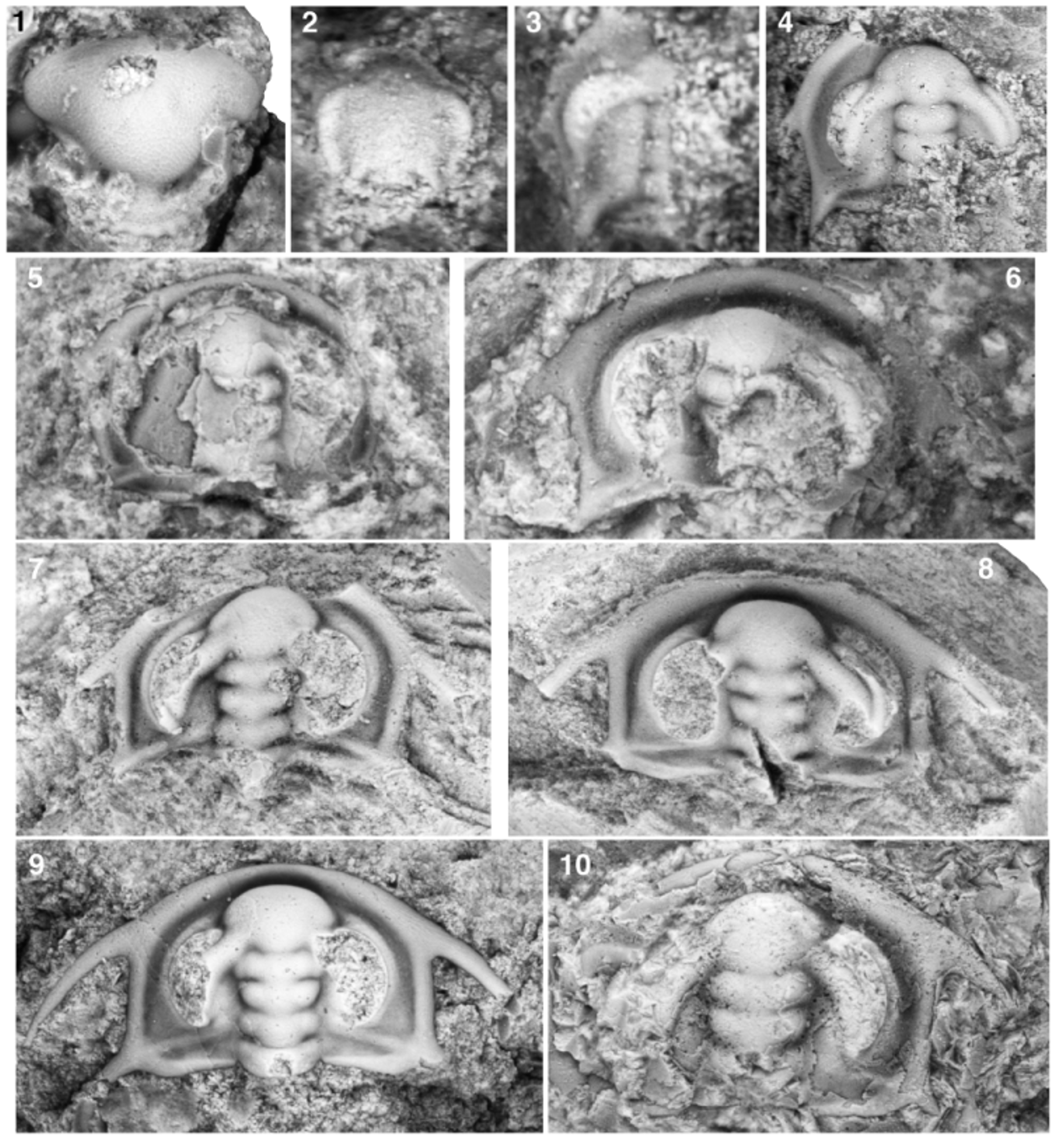
Figs 16 View FIGURE 16 , 17 View FIGURE 17
1972 Laudonia; Fritz, p. 27.
1992 Laudonia? sp. cf. Laudonia? mexicana (Lochman) ; Fritz, p. 11.
Diagnosis. Cephalon subquadrate in outline; genal spine bases located transversely opposite anterior-most point of S3 or posterior portion of LA (sag.); distal portion of posterior cephalic margin oriented anteriorly and inwards at 0° to 12° relative to an exsagittal line when traced toward base of genal spine. Posterior tip of ocular lobe transversely opposite lateral margin of L1.
Description (mature morphology, phase 5 of cephalic development; sagittal cephalic length> 5 mm)
Cephalon subquadrate in outline, width (tr.) at base of intergenal spines equal to or very slightly exceeds width (tr.) at base of genal spines. Cephalic length of largest known specimen estimated to have been approximately 17 mm (sag.). Proximal portion of posterior cephalic margin oriented weakly posteriorly by up to 10° relative to a transverse line when traced abaxially, flexing anteriorly at a rounded adgenal angle located approximately onethird distance along cephalic margin from axial furrow to base of genal spine, distal portion of posterior cephalic margin oriented anteriorly and inwards at 0° to 12° relative to an exsagittal line when traced toward base of genal spine. Genal spine rather broad-based, tapers along length; angle between distal portion of posterior cephalic margin and inner margin of proximal portion of spine approximately 64° to 74°, spines curving into more posterior orientation along their length; spine length estimated to be approximately three-quarters cephalic length (sag.; but spine tips not preserved on any specimen examined herein); spine base at contact with distal portion of posterior cephalic margin transversely opposite anterior-most point of S3 or posterior portion of LA (sag.). Distance (tr.) between genal spine bases approximately 133% to 144% of cephalic length (sag.). Intergenal spine slender, rather broad-based, angle between distal portion of posterior cephalic margin and outer margin of proximal portion of spine approximately 149° to 161°; spine length estimated to be approximately two-fifths of cephalic length (sag.; but spine tips not preserved on any specimen examined herein); spine base slightly distal to adgenal angle. Cephalic border defined by broad, trough-like border furrow interrupted only by intergenal ridge as it merges with border at base of intergenal spine; border furrow somewhat shallower and narrower along proximal portion of posterior cephalic border. Anterior cephalic border narrowest anterior to LA, broadens distally, width opposite junction of ocular lobes with LA approximately 15% length (sag.) of glabella; broadest portion of border located at confluence with genal spines; posterior border tapers to point adaxially, barely touches axial furrow at posteriormost portion of LO. Border well rounded dorsally anterior to LA, somewhat more broadly dorsally arched at and between bases of genal and intergenal spines. Preglabellar field absent; LA slopes anteriorly almost vertically into or slightly overhangs trough-like cephalic border furrow that is approximately as wide as anterior border at sagittal axis. Plectrum not developed. Glabella club-shaped in outline, most constricted (tr.) at L1, 90% to 93% of cephalic length (sag.), moderately dorsally convex (tr.), strongly dorsally arched (sag.), summit formed by posterior portion of LA and L3. Maximum width of LA averages 152% (range 142% to 170%) of basal glabellar width (tr.). Posterior margin of glabella convex posteriorly. SO transglabellar but deepest abaxially, abaxial end slightly anterior to adaxial end. LO subquadrate (transverse width across SO averages approximately 97% [range 94% to 100%] of basal glabellar width), length (exsag.) approximately 10% to 12% of glabellar length (sag.), very strongly dorsally convex (tr.). S1 transglabellar but deepest abaxially, approximately parallel to SO, slightly deeper than SO over axis. L1 weakly trapezoidal (narrowing slightly anteriorly), transverse width across S1 approximately 90% (range 83% to 97%) of basal glabellar width; length (exsag.) approximately 11% to 14% of glabellar length (sag.). S2 transglabellar but deepest abaxially and shallower than S1 over axis, connected to axial furrow, strongly arcuate (convex anteriorly) on either side of sagittal axis, adaxial portion slightly to well posterior to distal portion. L2 broadly V-shaped, lateral margins strongly diverging anteriorly, transverse width across S2 approximately 110% (range 101% to 122%) of basal glabellar width, distance (exsag.) between contact of S1 with axial furrow and contact of S2 with axial furrow approximately 11% (range 10% to 13%) of glabellar length (sag.). S3 transglabellar, oriented strongly anterolaterally away from axis until contact with ocular lobes where it contacts anteriorly converging axial furrow, the two together thus forming a strongly caret-shaped furrow on either side of sagittal axis, deepest at apex of each caret. L3 broadly M-shaped, lateral margins diverging anteriorly until contact with inner margin of ocular lobes, then converging anteriorly until contact with S3; transverse width of glabella at point of contact between axial furrow and inner margin of ocular lobes approximately 124% (range 116% to 138%) of basal glabellar width, distance (exsag.) between contact of S2 with axial furrow and contact of axial furrow with inner margin of ocular lobes approximately 4% of glabellar length (sag.). LA impinges into and anteriorly deflects anterior border furrow; transversely oblate in outline, anterior margin broadly rounded; maximum width (tr.) at or very slightly anterior to contact with outer margin of ocular lobes, length (sag.) approximately 67% (range 61% to 78%) of maximum width and 41% to 45% of glabellar length (sag.), maximum width (tr.) approximately 152% (range 143% to 170%) of basal glabellar width; moderately dorsally convex (tr.), strongly dorsally convex (sag.); prominently inflated relative to extraocular area, lateral margins clearly defined from anterior portion of extraocular area by axial furrow. Shallow, weakly arcuate (convex posteriorly) preocular furrow runs inwards and anteriorly from contact of LA with outer margin of ocular lobes; not incised over axis. Very shallow transocular furrow runs anterolaterally across ocular lobes from point of contact between S3 and axial furrow to preocular furrow, isolating ocular lobes from LA. Axis of LO often chipped, but preserves base of prominent axial node or short axial spine (Fig. 16.1, 16.9). Subdued ovoid lateral swellings on L1 and L2, very subdued lateral swellings on anterior portion of LO and sometimes on L3 (Fig. 16.9); lateral swellings on LO merge into interocular area, interrupting axial furrow. Axial furrow broad, deepest at lateral margins of L2 and L3. Ocular lobes divergent from exsagittal axis by approximately 26° (range 22° to 31°), crescentic; posterior tip transversely opposite mid- to anterior portion of lateral margin of L1 (more anteriorly located on larger specimens). Deep ocular furrow runs along length of ocular lobe slightly distal to midline so that inner band of ocular lobe is wider (tr.) than outer band; anteriorly, ocular furrow curves slightly outwards to merge into a “triple junction” of furrows at point where preocular furrow meets the axial furrow at the margin of LA, thus isolating outer band of ocular lobe from LA; posteriorly, ocular furrow curves slightly inwards to meet inner margin of ocular lobe so that posterior tip of ocular lobe is located on outer band. Inner and outer bands of ocular lobe each convex dorsally (tr.). Circumocular suture defines large visual surface (not preserved) that occupied much of length of outer wall of ocular lobe; portion of outer wall of ocular lobe below circumocular suture forms prominent, steep eye socle approximately one-third of estimated vertical height of visual surface (Fig. 16.7). Interocular area slopes down from inner margin of ocular lobe to axial furrow; approximately half width (tr.) of inner band of ocular lobe opposite S2. Extraocular area climbs out of broad border furrow then arches to produce a moderately vaulted transverse profile. In vicinity of ocular lobe, extraocular area abruptly changes slope to form a steep-walled, narrow-topped (tr.) extraocular platform upon which ocular lobe sits (Fig. 16.6, 16.7); extraocular platform subcrescentic in outline when viewed from above; anteriorly, extraocular platform narrows to a thin, subdued ridge that extends beyond anterior limit of ocular lobe, descending in vertical height and running parallel to lateral margin of LA then fading into anterior border furrow; posteriorly, extraocular platform terminates just posterior to posterior tip of ocular lobe. Prominent intergenal ridge extends from interocular area opposite lateral margin of L1 to posterior cephalic border at base of intergenal spine; ridge slightly broadens distally and merges smoothly into border, interrupting border furrow. Network of raised ridges forming walls of polygonal, mesh-like prosopon on dorsal surface of extraocular area, cephalic border furrow, cephalic border, intergenal ridge, bases of intergenal and genal spines, and sometimes interocular area, ocular lobes, and glabella (Fig. 16.2, 16.3, 16.5, 16.10, 16.11); polygons become more elongate (long axes parallel to outer wall of ocular lobe) on extraocular platform, more elongate (long axes parallel to axial furrow) on LA, and more elongate (long axes parallel to cephalic margin) on outer portion of distal posterior cephalic border and anterior cephalic border, often grading into bertillon-like pattern of anastomosing ridges on outermost cephalic border and genal and intergenal spine bases that in turn grades into subparallel lirae on sagittal portion of anterior cephalic border and distal portions of genal and intergenal spines (Fig. 16.1, 16.2, 16.9, 16.11). Central raised boss developed in some polygons (Fig. 16.11); central bosses developed into large, pustule-like structures in some polygons on distal portion of extraocular area, cephalic border furrow, intergenal ridge, posterior cephalic border (particularly on inner portions), and on base of intergenal spines.
Only known hypostome (Fig. 17.1) approximately 3.89 mm in sagittal length, moderately convex (tr., sag.), subtrapezoidal (widest anteriorly) in outline. Anterior lobe of middle body subtriangular, occupies approximately 75% of sagittal length of hypostome; anterior margin broadly curved; extends laterally onto broad, triangular anterior wings, tips of which are located approximately one-third distance along hypostomal length (traced from anterior to posterior). Posterior lobe of middle body narrow, <20% of hypostome length (sag.), transversely crescentic in outline, widest anteriorly, distinguished from anterior lobe over sagittal axis by broad, shallow furrow. Maculae relatively deep. Anterior border very narrow, rim-like; defined at least abaxially by shallow furrow (not preserved sagittally). Posterior border narrow, approximately 6% of hypostome length sagittally, weakly defined by shallow furrow. Six(?) pairs of small spines/nubbins project posterolaterally from margin of lateral and posterior border; abaxial-most three(?) pairs not well preserved, fourth and fifth pairs developed as short, stubby spines, sixth (innermost) pair nubbin-like and located close to sagittal axis. Network of raised ridges forming walls of polygonal, mesh-like prosopon on ventral surface of anterior and posterior lobes; central raised boss developed in some polygons. Rostral plate, thorax, and pygidium unknown.
Ontogeny. Specimens examined herein range from approximately 0.59 mm to approximately 17 mm in cephalic length (sag.). The smallest known cephalon (Fig. 17.2) lacks genal spines and is therefore assigned to prephase 3 of cephalic development; the size of the specimen is also consistent with that of pre-phase 3 cephala of other olenelloids. All larger specimens represent phases 3 to 5 of cephalic development. Specimens of sagittal cephalic length> 5 mm are essentially morphologically mature and have been described previously.
Pre-phase 3: Only known specimen approximately 0.59 mm in cephalic length (sag.; Fig. 17.2). Cephalon probably horseshoe-shaped in outline (although the anterolateral margins are worn and a more polygonal original outline cannot be ruled out); gently dorsally convex (tr. and sag.). Very short, stubby, triangular intergenal spines at posterolateral corners of cephalon; distance (tr.) between inner margins of intergenal spine bases approximately 46% of sagittal cephalic length. Anterior cephalic border broadly dorsally convex in cross-section, separated from LA by trough-like border furrow that is approximately as broad (sag.) as anterior cephalic border; border extends laterally around cephalic margin until it abuts and is truncated by outer margin of ocular lobes. Glabella occupies approximately 84% of cephalic length (sag.). Axial furrow not incised between ocular lobes, glabella not externally differentiated from shelf-like interocular area; glabellar furrows and interocular furrows similarly not expressed, giving cephalon an effaced appearance. LA clearly differentiated from frontal area as a slightly inflated, transversely oblate structure with a broad, bluntly rounded anterior margin; width (tr.) of LA at point where it contacts outer margin of ocular lobe slightly greater than distance (tr.) between inner margins of intergenal spine bases, LA therefore wider (tr.) than basal glabellar width. Crescentic ocular lobes diverge outwards and posteriorly from posterolateral margins of LA, then gently curve such that they are weakly convergent posteriorly, abut the cephalic margin at point of widest separation, then run confluent with margin posteriorly. Inner margin of each ocular lobe differentiated from interocular area by shallow furrow. Length of each ocular lobe approximately 80% of glabellar length (sag.). Maximum transverse distance between outer margins of ocular lobes approximately twice width (tr.) of LA where it contacts outer margins of ocular lobes. Dorsal surface of each ocular lobe convex upwards in cross-section; ocular furrow absent. Each ocular lobe tapers posteriorly and merges smoothly into a weak posterior ocular line that is oriented inwards and posteriorly and contacts abaxial portion of intergenal spine base. Posterolaterally oriented intergenal ridge runs from posterior portion of interocular area (presumably representing a pleural extension of L1) to posterolateral corner of cephalon where it merges onto intergenal spine. Axial nodes not present on glabella. Presence or absence of procranidial spines cannot be determined.
Phase 3: Initial morphology: Smallest known specimen approximately 0.9 mm in cephalic length (sag.; Fig. 17.3); only left side of cephalon is preserved. Cephalon horseshoe-shaped in outline. Posterior cephalic margin between bases of intergenal spines more-or-less transversely oriented. Intergenal spines project posterolaterally from posterolateral corners of cephalon; tips not preserved; distance (tr.) between inner margins of intergenal spine bases estimated to be approximately 58% of cephalic length (sag.). Anterior border gently rounded dorsally, sagittal length estimated to be approximately 14% of glabellar length (sag.), narrows laterally and posteriorly; border extends around lateral cephalic margin to base of intergenal spines. Tiny, thorn-like genal spine projects posterolaterally from lateral margin at widest point of cephalon (tr.), approximately transversely opposite midpoint of sagittal axis and posterior-most portion of lateral margin of L3. Slight geniculation of cephalic margin at anterolateral corner of cephalon where short procranidial spines project anteriorly, another geniculation of cephalic margin midway between base of procranidial spines and genal spines; cephalic margins between genal and intergenal spines convergent posteriorly. Glabella evenly tapers anteriorly from SO to S3; approximately 83% of cephalic length (sag.); defined by shallow axial furrow. SO, S1, S2, and S3 deepest abaxially, straight; SO oriented slightly anterolaterally when traced abaxially, all other glabellar furrows more-or-less transverse. Preocular furrow not present. LO probably rectangular or subtrapezoidal (widening slightly [tr.] anteriorly) in outline; sagittal length approximately 11% of glabellar length (sag.); lateral margins poorly differentiated from posterior cephalic border and posterior portion of interocular area; posterior margin convex posteriorly. L1, L2, and L3 approximately 16%, 19% and 19% of cephalic length (sag.), respectively; each slightly trapezoidal in outline (narrower anteriorly). LA transversely oblate, widest (tr.) at point of contact with outer margin of ocular lobes; clearly delimited from preglabellar field by break in slope; summit slightly lower than summit of ocular lobes. Maximum width (tr.) of LA slightly wider than basal glabellar width. Presence or absence of axial nodes on glabella cannot be determined. Ocular lobes crescentic in dorsal view, differentiated from interocular area by furrow; strongly divergent proximally, bending to run almost parallel with exsagittal axis; posterior tips more widely separated than anterior tips and located transversely opposite anterior-most lateral margin of L1; angle between exsagittal axis and line from posterior tip of ocular lobe to contact of inner margin of ocular lobe with lateral margin of LA approximately 41°; convex in cross-section (tr.); dorsal summit slightly higher than highest points of interocular area and anterior portion of glabella. Interocular area weakly dorsally arched (tr.); interocular furrows not visible. Intergenal ridge originates adjacent to lateral margin of L1, runs onto adaxial portion of intergenal spines; much weaker posterior ocular line extends to abaxial margin of intergenal spines. Very narrow (tr.), crescent-shaped extraocular area extends posteriorly to posterior ocular line; sagittally developed as broad, trough-like border furrow (or preglabellar field?) approximately as broad (sag.) as anterior cephalic border.
Subsequent morphological change: A slightly larger specimen (Fig. 17.4; sagittal glabellar length estimated to be approximately 1.66 mm) is also incompletely preserved, but represents a cephalon in either very late phase 3 or very early phase 4 of development and, by comparison to the previous specimen, offers insight into some of the morphological changes that occurred during phase 3 of cephalic development. On the glabella, LA proportionally elongated (sag.) to occupy more than 36% of estimated sagittal glabellar length and inflated in dorsal relief. The sagittal elongation of LA resulted in a slight posterior deflection of the sagittal portions of S3, S2, and S1, so that they became oriented slightly anterolaterally when traced abaxially. L3 proportionally widened slightly, so that its anterolateral corners contacted the inner margin of the ocular lobes—S3 therefore became very slightly caretshaped on either side of the sagittal axis—and the glabella became lollipop-shaped, seemingly almost parallelsided between S3 and SO. (Incomplete preservation of the posterior portion of the glabella means that the widths [tr.] of the glabella across SO, S1, and S2 are uncertain, leading to ambiguity as to whether this specimen had entered phase 4 of cephalic development.) The preocular and transocular furrows became visible, albeit very faintly. No axial structures are evident on L2, L3, or LA on this specimen (L1 and LO are not preserved). The posterior tips of the ocular lobes became more distinctly differentiated from the (faint) posterior ocular line, and are located transversely opposite the anterior portion of the lateral margin of L1. An ocular furrow developed. The extraocular area proportionally widened (tr.), with a corresponding proportional elongation (tr.) of the posterior cephalic margin between the axial furrow and the base of the intergenal spine and an increased separation (tr.) between the outer margin of the ocular lobe and the cephalic border. The intergenal spine bases, which were initially located more-or-less directly behind (exsag.) the posterior tips of the ocular lobes on the smaller phase 3 cephalon, migrated to positions posterior to and almost outside the abaxial-most points of the ocular lobes. The genal spine proportionally elongated (although the tip is not preserved, so the full length of the spine is unknown). The intergenal spine also proportionally lengthened into an elongate triangular structure almost as long as the combined length (exsag.) of L2 and L3. The anterior portion of the cephalon is missing on this specimen, and neither the presence/absence of procranidial spines, nor the relative breadth of the (seemingly wide) anterior border furrow/preglabellar field, can be reliably determined. A weak geniculation is still evident in the cephalic margin slightly anterior to the genal spines.
Phase 4: The smallest known specimen that undoubtedly represents phase 4 of cephalic development has an estimated sagittal cephalic length of approximately 1.98 mm (Fig. 17.5). The specimen is poorly preserved, but is assigned to early phase 4 of cephalic development based on the fact that L3 is slightly wider (tr.) than L2, indicating that the pronounced lateral expansion of L3 relative to LA and L2 that defines entry into phase 4 had begun. LA continued its proportional elongation (sag.) from phase 3, and S3 is more strongly caret-shaped on either side of the sagittal axis. A weak intergenal angle is developed along the posterior cephalic margin approximately midway between the axial furrow and the inner margin of the intergenal spine base. No procranidial spines are evident.
Two slightly larger specimens (Fig. 17.6, 17.7; sagittal glabellar lengths estimated to be approximately 1.73 mm and 2.09 mm, respectively) represent either late phase 4 or earliest phase 5 of cephalic development—their precise phase assignment is ambiguous due to incomplete preservation of the glabella (Fig. 17.6) and slight lateral shearing (Fig. 17.7) that, in both cases, renders the precise form of L3 unclear. Nevertheless, through comparison with smaller specimens, these specimens reveal the nature of morphological change achieved during phase 4 of cephalic development. On the glabella, there was a pronounced lateral widening (tr.) of L3 relative to L2, resulting in an increased area of contact between the anterolateral margins of L3 and the inner margins of the ocular lobes. There was also a proportional lateral widening of LO and L1 relative to L2, so that the glabella became narrowest (tr.) at L2. LA continued to proportionally elongate (sag.), so that by the end of phase 4 it is estimated to have occupied approximately 40% of sagittal glabellar length and was separated from the anterior cephalic border by a trough-like border furrow. The larger specimen (Fig. 17.7) shows weak lateral swellings on L2 and L3, and the chipped base of an occipital node or spine. By late phase 4 the posterior ocular line was very faint, running close to and more-or-less parallel with the intergenal ridge for much of its length. The genal spines proportionally elongated into slender spines (although their full length is unknown) and migrated anteriorly (by the end of phase 4 the genal spine bases were located transversely opposite the midlength of the lateral margins of L3). The extraocular area continued to proportionally widen, especially posteriorly. This resulted in the base of the intergenal spines migrating to a position outside (exsag.) the distal-most part of the ocular lobe, the distal portions of the posterior cephalic margins rotating from being anteriorly divergent into a more nearly parallel-sided orientation, and the intergenal ridge becoming more transversely oriented. The intergenal spines lengthened (although their full length is unknown) and became more strongly laterally flared. The anterior cephalic border proportionally broadened relative to the exsagittal length of L1 and L2. A narrow extraocular platform was well developed around each ocular lobe by the end of phase 4.
Phase 5: Several specimens ranging from approximately 2.40 mm to approximately 3.34 mm in sagittal cephalic length are assigned to phase 5 of cephalic development based on exhibiting an L2 that widens (tr.) anteriorly (with the glabella being narrowest [tr.] at S1), indicating that L2 had initiated the pronounced lateral widening (tr.) that defines entry into that phase (Fig. 17.8–17.10). However, these specimens differ from the mature morphology (sagittal cephalic length> 5 mm; described previously) in several respects, and thus reveal several ontogenetic changes that occurred during early phase 5 of cephalic development. During early phase 5 of development, LA continued to proportionally elongate (sag.), occupying almost 40% of sagittal glabellar length upon entry into phase 5 and typically approximately 43% of sagittal glabellar length at morphological maturity (Fig. 18.1). As a result, glabellar furrows SO, S1, and especially S2 and S3 became deflected into progressively more strongly posterolaterally directed orientation (when traced adaxially). During phase 5, LA progressively impinged into the anterior border furrow as its elongation resulted in the glabella occupying a greater proportion of sagittal cephalic length (Fig. 18.2). The entire glabella proportionally laterally widened (tr.) relative to its length; the area of contact between the anterolateral margins of L3 and the inner margins of the ocular lobes therefore continued to increase, and the ocular lobes therefore became less strongly divergent (Fig. 18.3). The posterior ocular line became obsolete. There was continued proportional elongation and anterior migration of the genal spines (Fig. 18.4). The anterior cephalic border continued to proportionally broaden, especially near the genal spine bases. The distal portions of the posterior cephalic margins continued to rotate into a nearly parallel and sometimes slightly anteriorly convergent configuration, although with much variation (Fig. 18.5; contrast Fig. 17.8, 17.9, and 17.10). There is accordingly considerable variation in the width (tr.) of the extraocular area, especially anteriorly (Fig. 18.6).
Etymology. Named for the subquadrate cephalic outline.
Holotype. GSC 137494 (Fig. 16.1), from GSC locality 95191, Cassiar Mountains, British Columbia.
Other material. Quantitative morphometric and/or qualitative descriptive data were recorded from first-hand examination of the holotype (see previous) and the following 23 cephala: three additional cephala from GSC locality 95191 ( GSC 137496, GSC 137499, and GSC 137500); nine cephala from GSC locality 82782 (including GSC 137495, GSC 137497, GSC 137498, and GSC 137501); five cephala from ICS-1336 (including FMNH PE 58487 to FMNH PE 58489); and six cephala from LACMIP locality 17053 (LACMIP 12539, LACMIP 12540, LACMIP 12541, LACMIP 12543, LACMIP 12552, and LACMIP 12554). One hypostome from LACMIP locality 17053 is also assigned to this species.
Occurrence. CANADA: Cassiar Mountains, British Columbia: GSC locality 95191, from a thin, wavybedded limestone 18.75 metres above the base of Unit 4 of the Rosella Formation, in Section 2 of Fritz (1978, textfig. 3.1a), on the north bank of the Dease River, approximately 2.7 km ( 1.7 miles) east of McDame. Cariboo Mountains, British Columbia: GSC locality 82782, from a platy, thin-bedded, bioclastic limestone and interbedded micaceous shale 51.5 metres above the base of the middle member of the Mural Formation in a measured section between Come Creek and Ptarmigan Creek (probably corresponding to locality 7 of Young [1972, fig. 1]). U.S.A.: Montezuma Range, Esmeralda County , Nevada: ICS-1336, bioclastic limestone from an unconstrained stratigraphic position in the lower Dyeran portion of the middle member of the Poleta Formation, IS-4 section of Hollingsworth (1999), Indian Springs Canyon. Slate Ridge, Esmeralda County, Nevada: LACMIP locality 17053, light grey packstone approximately 150 metres above the base of the section (approximately 134 metres above the base and 55 metres below the top of the middle member of the Poleta Formation), in the Gold Point section of Bohach (1997, text-fig. 62, p. 374).
Discussion. Lochmanolenellus subquadratus is easily distinguished from Lo. primus and Lo. pentagonalis by its subquadrate rather than pentagonal cephalic outline (Figs 9.1, 9.2, 18.8), and by its more anteriorly positioned genal spine bases (Fig. 9.3). Lochmanolenellus subquadratus is similar to Lo. trapezoidalis in terms of degree of genal spine advancement (Fig. 9.3), but differs from that species in several other respects. First, the ocular lobes of Lo. subquadratus are longer (with the posterior tips being located transversely opposite the lateral margin of L1) than those of Lo. trapezoidalis (with the posterior tips being located transversely opposite the distal tip of S1 or lateral margin of L2; Fig. 18.7). Second, the distal portion of the posterior cephalic border is oriented almost directly anteriorly or very slightly inwards when traced towards the base of the genal spine on morphologically mature cephala of Lo. subquadratus (resulting in a more-or-less subquadrate cephalic outline), but is consistently oriented more strongly inwards on mature cephala of Lo. trapezoidalis (resulting in a distinctly more trapezoidal cephalic outline; Figs 9.1, 18.8). Third, the intergenal spines of Lo. subquadratus flare outwards (relative to the orientation of the distal portion of the posterior cephalic margin) more strongly than on Lo. trapezoidalis (Fig. 9.5). Lochmanolenellus subquadratus and Lo. trapezoidalis share many similar ontogenetic trends of morphological change. However, in contrast to Lo. trapezoidalis , the ocular lobes of Lo. subquadratus did not appreciably proportionally shorten (relative to glabellar length) during phase 5 of cephalic development, but remained approximately half as long as the glabella (Fig. 18.7) with their posterior tips remaining transversely opposite the lateral margins of L1. Indeed, for cephala larger than 1 mm in sagittal glabellar length, the allometric relationship between (log-transformed) ocular lobe length and (log-transformed) glabellar length is significantly different between the two species (for Lo. subquadratus [n = 8], RMA slope = 0.933; for Lo. trapezoidalis [n = 17], RMA slope = 0.779; slopes significantly different at 95% confidence; analysis performed using smatr package in R [ Warton et al., 2012; R Core Team, 2013]).
The hypostomes of Lo. subquadratus and Lo. trapezoidalis are also similar in most respects. The innermost pair of spines on the posterior margin of the hypostome is located much closer to the sagittal axis on Lo. subquadratus than on Lo. trapezoidalis , although this may reflect an ontogenetic difference because the only known hypostome of Lo. subquadratus is much smaller than the smallest known hypostome of Lo. trapezoidalis .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Olenellina |
|
Family |
|
|
Genus |