
Petropedetes vulpiae, Barej, Michael F., Rödel, Mark-Oliver, Gonwouo, Legrand Nono, Pauwels, Olivier S. G., Böhme, Wolfgang & Schmitz, Andreas, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.193288 |
|
DOI |
https://doi.org/10.5281/zenodo.6199588 |
|
persistent identifier |
https://treatment.plazi.org/id/B479090F-FFC3-FF86-D7B4-FE4641940CE0 |
|
treatment provided by |
Plazi |
|
scientific name |
Petropedetes vulpiae |
| status |
sp. nov. |
Petropedetes vulpiae View in CoL sp. nov.
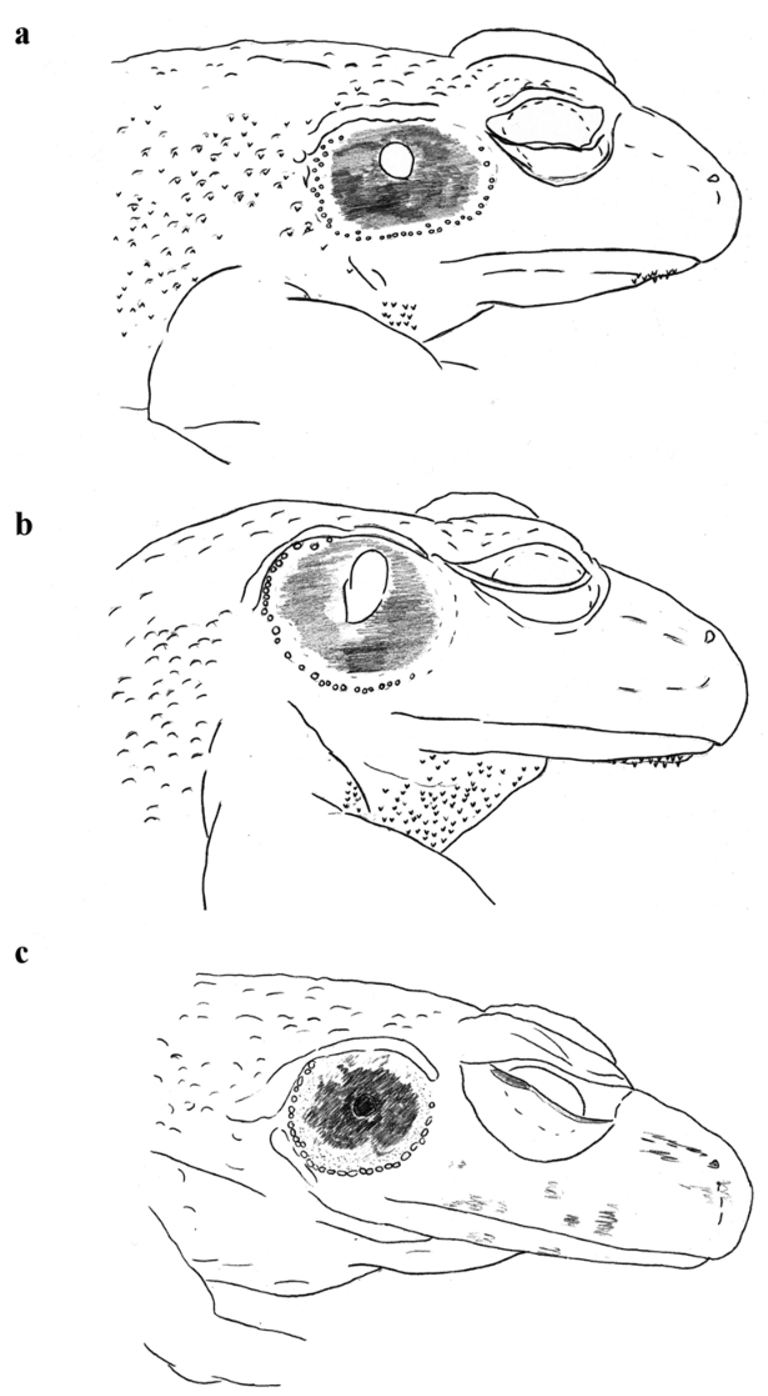
Figs. 1 View FIGURE 1 b, 2b, 4, 6a, 7b, 7c, 7d, 7e
Holotype. ZFMK 81623 (adult male), Cameroon, Mt. Nlonako, Ekomtolo, 4°51’N, 9°54’E, app. 450 m a.s.l., 12.-16.V.2003, coll. M. Plath.
Paratypes. IRSNB-KBIN 1987 (male) Cameroon, foot of Rumpi Hills, Big Massaka, 4°41’44N, 09°17’ 31.4E, app. 315 m a.s.l., 8.VII.2009, coll. A. Schmitz & M.F. Barej; MHNG 2713.1-2713.3 ( 3 males), same data as IRSNB-KBIN 1987; MHNG 2713.6 (female), Cameroon, Mt. Nlonako, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., 7.II.2003, coll. M. Plath; ZFMK 75588 (male), Cameroon, Mt. Kupe, Nyasoso, 4°49’42N, 9°40’55E, app. 920 m a.s.l., 12.II.2000, coll. H.-W. Herrmann & A. Schmitz; ZFMK 81167 (male), Cameroon, Mt. Nlonako, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., 17.VIII.2000, coll. M. Solbach; ZFMK 81553 (male), Cameroon, Mt. Nlonako, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., 13.I.2001, coll. M. Plath; ZFMK 81554 (female), Cameroon, Mt. Nlonako, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., no date, coll. M. Plath; ZFMK 81711 (male), Cameroon, Mt. Nloanko, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., 14.V.2003, coll. M. Plath; ZFMK 88863 (male) same data as IRSNB-KBIN 1987; ZMB 73690 (female), Cameroon, Mt. Nlonako, Ekomtolo, 4°49’49N, 09°54’43E, app. 450 m a.s.l., 31.III / 4.IV.2001, coll. H.-W. Herrmann et al.; ZMB 73691-92 ( 2 males), same data as IRSNB-KBIN 1987.
Additional material examined. IRSNB-KBIN 13.841 (male), Gabon, Estuaire Province, Kango Department, 15 north of Alen-Nkoma, right of route L107, 2004, coll. O.S.G. Pauwels; IRSNB-KBIN 13.845 (female), 13.846 (male), Gabon, La Ngounié Province, Itsila, July 2004, coll. O.S.G. Pauwels; IRSNB-KBIN 14.932-933 ( 2 males) 14.934 (female), Gabon, Province de l’Estuaire, Kinguélé, app. 80 m a.s.l., IV.2001, coll. O.S.G. Pauwels; IRSNB-KBIN 14.935 (male), Gabon, Province du Woleu-Ntem, Song, app. 360 m a.s.l., 25.VI.2001, coll. O.S.G. Pauwels; MHNG 684.7 ( 2 males), Cameroon, Efulen, Kribi, no date, coll. Rosenberg; MHNG 917.70-71 ( 2 females), Cameroon, Ebolowa, Nkoétyé, 1.I.1955, coll. J.-L. Perret; MHNG 1033.67 (male), Cameroon, Makai, river Ngopi, 1960, coll. J.-L. Perret; MHNG 1521.80 (male), 1521.82 (male), 1521.84 (female), Cameroon, Sakbayémé, 23.X.1972, coll. J.-L. Perret; MHNG 1521.87 (male), 1521.88-90 ( 3 females), Cameroon, Kala, X.1972, coll. J.-L. Perret; MHNG 1521.92-93 ( 2 females), Cameroon, Ebolowa, Njazeng, 22.IV.1972, coll. J.-L. Perret; MHNG 1521.94 (male), Cameroon, Ototomo, 30.XII.1968, coll. J.-L. Amiet; MHNG 1521.95 (male), Cameroon, Maholé, Mungo, 28.III.1973, coll. J.-L. Perret; MHNG 2713.4 (female), Cameroon, Mt. Kupe, Nyasoso app. 950 m a.s.l., 5.VII.2009, coll. A Schmitz & M.F. Barej; ZFMK 19582 (male), 19585 (male), 19587 (male), Cameroon, Fako, Mt. Kupe, 5.-6.XII.1976, coll. U. Joger & J. Joger; ZFMK 69202 (male), 69209 (female), Cameroon, Mt. Nlonako, above Nguengue, app. 1100–1200 m a.s.l., 23.XI – 6.XII.1998, coll. ALSCO-Expedition; ZFMK 88860 (male), data as MHNG 2713.4; ZFMK 88861, ZFMK 88862 ( 2 males), 6.VII.2009, other data as MHNG 2713.4; ZMB 73726 (male), Nigeria, Mbe Mts., X.2005, coll. A. Onadeko; ZMB 73893 (male), Gabon, Province du Woleu-Ntem, Tchimbélé, app. 469 m a.s.l., 10.IV.2001, coll. O.S.G. Pauwels; ZMB 73894 (female) same data as IRSNB- KBIN 14.935.
Diagnosis. Medium sized Petropedetes ; compact body shape; tympanum usually flattened on the upper and lower border; tympanum larger than diameter of eye in males ( Fig. 1 View FIGURE 1 b), smaller in females; characters of breeding males: tympanic papilla present (broad, fleshy), papilla closer to the centre than the upper border; forearm hypertrophy well developed; carpal spike present; spinosities on throat, forearms and on almost every wart on flanks and dorsum, even around the tympanum ( Fig. 6 View FIGURE 6 a); femoral glands large, very prominent; webbing rudimentary.
Description of the holotype. The holotype is an adult male in breeding condition, robust body shape; 40.4 mm SUL; snout very short, rounded in lateral view; head width 17.4 mm; eye diameter 6.7 mm; interorbital distance 3.9 mm; naris closer to snout ( 2.2 mm) than to eye ( 4.5 mm); skin on throat with spinosities ( Fig. 6 View FIGURE 6 a); femur length 31.4 mm; tibia length 25.7 mm; foot (incl. longest toe) 31.4 mm; femoral glands large (length x width: 6.2 x 3.3 mm / 6.4 x 3.4 mm); relation femur length and femoral gland length 0.28; spinosity well developed on upper forearms and flanks; webbing rudimentary; coloration of preserved specimen: dorsum pale with dark spots (approx. 2–2.5 mm diameter); hind limbs marbled in dark and bright colours, but with distinct cross bars; throat dark; ventrum whitish with minuscule black speckles; femoral glands almost without darker pigments.
Variation. Medium sized Petropedetes with compact body; females slightly larger than males (SUL in males: 33.4–43.4 mm, females: 21.9–46.5 mm); mean head width in males 41% of SUL, in females 39%; snout in lateral view generally rounded; canthus rostralis sharp; loreal region concave; eye diameter about 1.6 times eye-narial distance; nose closer to snout tip than to eye; tympanum distinct, larger than the eye in males, smaller in females (tympanum / eye in males: 0.86–1.13, in females: 0.48–0.68); tympanum usually flattened on upper and lower border; tympanic papilla closer to centre than to upper border; tympanic papilla broad and fleshy; tympanum surrounded by minuscule white warts; supratympanic fold distinct; fingers slender, with Tshaped fingertips; relative length of fingers: III> IV> II> I; manual subarticular tubercles single; manual webbing absent; palmar tubercle and thenar tubercle present; forearm hypertrophy well developed in males; carpal spikes present in males; spinosities on throat, forearms, as well as on almost every wart on flanks and dorsum; dorsal tegument predominantly uniform with small regularly scattered warts, a few larger, partly linearly arranged warts present; granules on flanks larger than on dorsum; structure of ventral skin smooth; mean femur length in males 53% of SUL, in females about 51%; mean tibia length in both sexes 60% of SUL; mean foot length in both sexes 76% of SUL; upper hind limbs of moderate width, lower hind limbs slender; femoral glands more prominent in males, in females sometimes even indistinct, gland values of the two sexes overlap, can therefore be misleading without the relation of tympanum to eye diameter (femoral gland /femur length in males: 0.16–0.40, in females: 0.21–0.29); relative length of toes: IV> III> V> II> I; webbing rudimentary: 1 (1), 2 (1-1), 3 (2-2), 4 (3-3), 5 (2).
Coloration. In life dorsum marbled in brown, green and whitish; larger warts on dorsum are usually dark coloured; iris whitish-golden; hind limbs with large dark spots, similar to transverse bars, large transverse bars are divided by thin light lines, large bars and thin light thin lines may continue on the lower leg ( Fig. 1 View FIGURE 1 b); throat in most specimens dark, otherwise whitish with dark pigments; belly usually whitish; femoral glands yellowish or orange. Coloration in preservation as in life, but pale; dorsum generally brown; belly whitish with minuscule black speckles (recognizable only under microscope).
Natural history. Petropedetes vulpiae sp. nov. is part of the Central African forest fauna ( Amiet 2006). Frétey & Blanc (2001) find the species close to a stream in forests and gallery forests. P. vulpiae sp. nov. is, just like P. aff. parkeri (see below), predominantly active at dawn and only becomes active during day when the climate is very humid ( Amiet 1989). Gabonese specimens were collected during midday, but also at night in small streams in primary forest. Specimens from Itsila ( Gabon) were caught on mossy rocks in the middle of a cascade at the border between Gabon and the Republic of the Congo ( Fig. 7 View FIGURE 7 b). Specimens from Alen- Nkoma ( Gabon) avoided jumping in fast running water while escaping, those who fell in the water were quickly exhausted (O.S.G. Pauwels, pers. obs.). P. vulpiae sp. nov. and P. aff. parkeri occur in sympatry and may share a similar biology. However, Amiet (1983) believes that P. v u l p i a e sp. nov. may occur on less steep surfaces than P. aff. parkeri . Perret & Mertens (1957) find specimens in a cave with a small current, where they hid in small holes between rocks. The authors assumed the species to be very common in its biotopes. The clutch of some ten eggs is deposited on moist surfaces on rocks ( Amiet 1983, 1989). Males seem to guard the eggs at night ( Amiet 1991). Lawson’s (1993) clutch observations seem to be based on P. j o h n s t o n i (see above). The call of P. vulpiae sp. nov. has been described as a gentle “douc-douc”, which is emitted by the males from hiding places within rock cavities ( Amiet 1973). The species shows a vocal annual activity cycle type II sensu Amiet (2006); a continuous cycle with one less intensive period at the end of the big rainy season and beginning of the following dry season. Mertens (1968) reports on mites in the vent region of several specimens in this species and Vaucher & Durette-Desset (1980) describe a member of the parasitic Spiruroidea from a specimen collected from Kala.
Distribution. Petropedetes vulpiae sp. nov. occurs from eastern Nigeria to southern Gabon ( Fig. 4 View FIGURE 4 ). The species is known from lowland localities, but also reaches altitudes above 1000 m (e.g. Mertens 1965; Plath et al. 2004). Lawson (1993) mentions a specimen from 1769 m from a locality in the Rumpi Hills, but remarked that it has been assigned to this species with reservation. The species is known from localities in Cameroon ( Boulenger 1906; Mertens 1965, 1968), Equatorial Guinea ( Perret 1984; de la Riva 1994; Lasso et al. 2002), and Gabon ( Frétey & Blanc 2001; Burger et al. 2006a, 2006b; Pauwels & Rödel 2007). It is also present in northern Republic of Congo, as specimens have been found on both sides of a cascade of the border between Gabon and the Republic of Congo (O.S.G. Pauwels, pers. obs.).
Genetics. The genetic comparison for the uncorrected p-distances of the mitochondrial 16S rRNAfragment (Table 4) gave the following results for Petropedetes vulpiae sp. nov.: interspecific comparison between P. vulpiae sp. nov. and all other Central African Petropedetes taxa ranged between 3.68%-9.53%, while the intraspecific variation within P. vulpiae was much lower at 0.00%-0.20% (N= 6). Specimens of P. vulpiae sp. nov. from Big Massaka perfectly correspond gentically to the holotype from Ekomtolo (Table 4) and are therefore included in the type material.
Systematic remarks. This species has long been considered to be P. newtonii . The taxonomic confusion concerning P. johnstoni and P. newtonii has been elucidated above, outlining that the “real” P. newtonii is a synonym of P. j o h n s t o n i. Petropedetes vulpiae sp. nov. shares the rudimentary development of its webbing with P. parkeri , P. johnstoni and P. euskircheni sp. nov. Hence, the new species is distinct from species with developed webbing ( P. p a l m i p e s and P. p e r re t i, both fully webbed; P. c a m e ro n e n s i s and P. juliawurstnerae sp.
nov., both half-webbed). The presence of a distinct tympanum and a tympanal papilla separates P. vulpiae sp. nov. from P. cameronensis and P. palmipes , who miss these characters.
Petropedetes vulpiae sp. nov. can be differentiated from P. johnstoni : by a) the size of the tympanum: the tympanum is smaller than the eye diameter in P. johnstoni , while it surpasses eye size in P. vulpiae sp. nov. (tympanum diameter / eye diameter in males: N vulpiae = 35, N johnstoni = 7, p <0.001); females of both species possess tympana smaller than the eye, but they are still larger in P. vulpiae sp. nov. (tympanum diameter / eye diameter in females: N vulpiae = 16, N johnstoni = 4, p <0.01); b) the size of femoral glands: the femoral glands are more developed in male P. johnstoni than in P. v u l p i a e sp. nov. (femoral gland length / femur length in males: N vulpiae = 35, N johnstoni = 7, p <0.05); there is no significant difference in gland size in females of both species ( p = 0.27).
Petropedetes vulpiae sp. nov. differs from P. parkeri and P. euskircheni sp. nov. by: a) the body size, P. vulpiae sp. nov. being smaller than P. p a r k e r i (SUL in males: N vulpiae = 35, N parkeri = 11, p <0.001; in females: N vulpiae = 16, N parkeri = 7, p <0.01) and P. euskircheni sp. nov. (see below in the species description of P. euskircheni sp. nov.); b) femoral gland size, the femoral gland is larger in males and females of P. vulpiae sp. nov. than in P. parkeri (femoral gland length / femur length in male: N vulpiae = 35, N parkeri = 11, p <0.001; in females: N vulpiae = 16, N parkeri = 7, p <0.001) and P. euskircheni sp. nov. (see below); the femoral glands of male P. parkeri can reach almost the size of females in P. vulpiae sp. nov.; in this case the femoral glands in male P. parkeri are more prominent and the tympanum is bigger than in female P. v u l p i a e sp. nov.); c) the tibia / SUL ratio is higher in both sexes of P. v u l p i a e sp. nov. (in males: N vulpiae = 35, N parkeri = 11, p <0.001; in females: N vulpiae = 16, N parkeri = 7, p <0.01); d) the shape of the tympanum, males of P. parkeri have a rounder tympanum than P. vulpiae sp. nov., it can be flattened in P. euskircheni sp. nov., but less than in P. v u l p i a e sp. nov. ( Fig. 6 View FIGURE 6 a).
Amiet (1987) mentions small morphological differences between P. vulpiae sp. nov. populations in Cameroon. Even more striking are differences in the size of the femoral gland between populations from northern Gabon and specimens from the type locality (own data). Unfortunately, we could not compare these populations genetically. Specimens from localities inbetween the type locality and Gabon exhibited intermediate stages in gland size. At the moment we regard all these populations as conspecific, however, their status should be reinvestigated when tissue of specimens from southern Cameroon and Gabon become available.
Etymology. The specific epithet vulpiae is the genitive of the latinised matronym of Dr. Christine Fuchs (in latin: Vulpes), Institut für Mikroskopische Anatomie und Neurobiologie der Universitätsmedizin der Johannes Gutenberg-Universität Mainz ( Germany), to whom we dedicate this new species, in recognition of her important contributions on globin research in African clawed frogs and her full and continuing support of A.S.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |