
Oswaldella occulta, Cantero & Molinero, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4457.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:463BBDB3-CBD9-43E3-9520-CDE336BC10ED |
|
DOI |
https://doi.org/10.5281/zenodo.5960858 |
|
persistent identifier |
https://treatment.plazi.org/id/B646223D-FFCA-D32D-FF24-113CFE4F3238 |
|
treatment provided by |
Plazi |
|
scientific name |
Oswaldella occulta |
| status |
sp. nov. |
Oswaldella occulta View in CoL sp. nov.
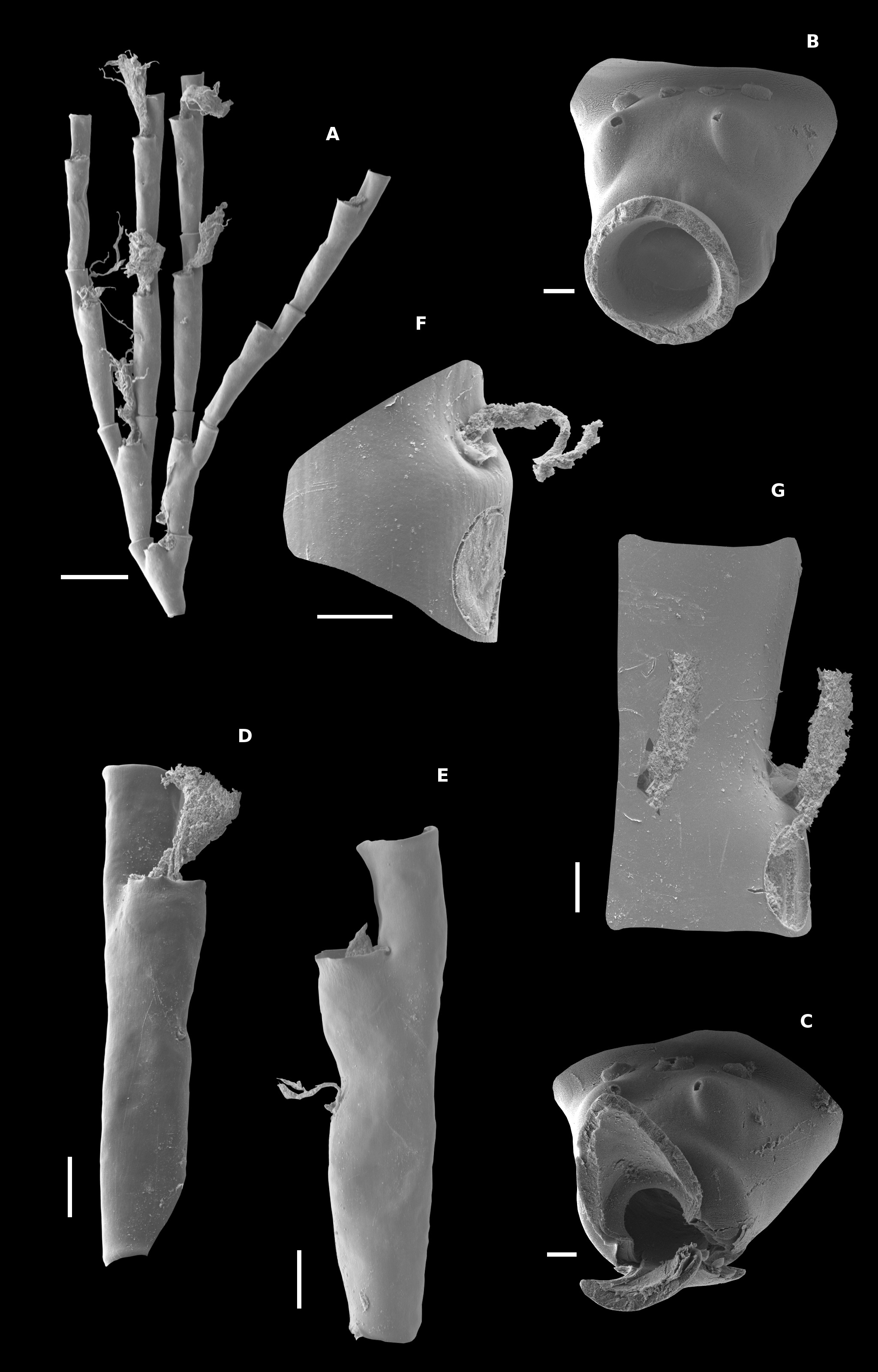
( Figs 7–8 View FIGURE 7 View FIGURE 8 )
Oswaldella sp. GOnzález MOlinerO & Peña CanTerO, 2015: 422, figs 15, 16P, 18H, 20H; Peña CanTerO, 2017: 60, fig. 19C.
Material examined. From the type series of Oswaldella terranovae: British Antarctic Terra Nova Expedition 1910–1913, Stn 220, 3 January 1912, off Cape Adare, Ross Sea, 82 – 92 m, several stems up to 120 mm high (NHM 1929.10.28.171), one slide with hydrocladia (1929.10.28.171, labelled n° 4), and four stems up to 130 mm high (NHM 1929.10.28.172). A 57-mm-long, basally broken stem from the material with the collection number NHM 1929.10.28.172 is designated as holotype (all the remaining material is paratype).
Other material examined: New Zealand Antarctic Expedition TAN0402, Stn 178, Tangaroa , 27 February 2004, 71°31’89’’–71°32’16’’S, 171°18’31’’–171°18’70’’E (Cape Adare, Ross Sea), 343–348 m (identified as Oswaldella sp. by González Molinero & Peña Cantero 2015).
Description. Stems at least 130 mm high, unbranched and slightly polysiphonic basally. Stem thin, yellowbrown, divided into internodes separated by strongly marked nodes. From one to three apophyses per internode. Angle between cauline apophyses and stem c. 45°. Cauline apophyses with two axillary nematophores, each emerging through a simple hole in perisarc ( Fig. 7B View FIGURE 7 ); no ‘mamelons’ present. Apophyses short, distinctly separated from hydrocladia.
Hydrocladia much branched ( Fig. 7A View FIGURE 7 ), with up to six third-order hydrocladia, three on each secondary hydrocladium (in one case with four tertiary hydrocladia on a secondary hydrocladium); occasionally, a short fourth-order hydrocladium was present. Tertiary hydrocladia placed lower than secondary ones. Hydrocladia divided into internodes, barely decreasing distally in size; e.g. 1000–930 µm in length and 280–270 µm in diameter under hydrotheca, in 1 st and 8th unforked internodes of secondary hydrocladium, respectively. First hydrocladial internode bifurcated ( Fig. 7A View FIGURE 7 ), with two similar prongs, distinctly separated from first internode of secondary hydrocladia. Mesial-inferior nematophore emerging from marked swelling at proximal third of internode ( Figs 7C– E View FIGURE 7 , 8A View FIGURE 8 ) and provided with a prominent nematotheca placed on frontal side of swelling ( Figs 7C–F View FIGURE 7 , 8 View FIGURE 8 ); usually nematotheca abcauline wall distinctly directed outwards. Hydrotheca placed on distal half of internode ( Figs 7A, C–E View FIGURE 7 , 8 View FIGURE 8 ). Hydrotheca low, about as high as wide. Abcauline hydrothecal wall straight, directed abcaudally; length slightly increasing along hydrocladium (e.g. 210 µm in 1 st internode of secondary hydrocladium and 240 µm in 7th internode). Perisarc of hydrotheca distinctly thickened ( Fig. 7C View FIGURE 7 ). Hydrothecal aperture circular ( Fig. 7C View FIGURE 7 ), 210–200 µm in diameter, roughly perpendicular to longitudinal axis of internode ( Figs 7D–E View FIGURE 7 , 8 View FIGURE 8 ); rim even ( Figs 7C–E View FIGURE 7 , 8 View FIGURE 8 ). In frontal view, hydrotheca usually jar-shaped, with a swollen basal half and a narrower distal one, but markedly widening again at aperture, producing a sort of narrow neck with flared rim ( Fig. 7C, E View FIGURE 7 ).
Gonothecae unknown.
Cnidome composed of microbasic mastigophores in two size groups: larger, range 11–15 x 3.5–4, mean 12.8 ± 1.2 x 3.8 ± 0.3 (n = 14); smaller, range 6– 7 x 2.
Remarks. The material of Oswaldella occulta sp. nov. was in the material assigned to O. antarctica ( Jäderholm, 1904) by Totton (1930) and later described as O. terranovae by Peña Cantero & Vervoort (1996).
On two occasions a secondary hydrocladium forms a third-order hydrocladium in its first and third internodes, but not in the second one (i.e. the second internode is unforked).
The species seems to have stems slightly polysiphonic only basally. Most of the stems available are deprived of the basal part and it was not possible to ascertain if they were polysiphonic. There are, however, hints indicating that the species can develop a sort of polysiphonic condition. Among the material examined, there is a stem, 105 mm high, provided with the basal part. Its first 13 mm have a few creeping stolons originating from cauline apophyses, some of them running downwards and acting as hydrorhizal stolons. In addition, some of the basally broken stems also have creeping stolons originating from cauline apophyses, although not in their most basal part. It is therefore highly likely that O. occulta sp. nov. can develop stems with a slightly polysiphonic basal part.
Oswaldella occulta sp. nov. appears to have unbranched stems, although the issue is as yet unresolved. The available material, however, points to the unbranched condition. The species seems to develop colonies with tuft appearance, with several unbranched stems growing together. In the available material there are some stems apparently branched, but the ‘branches’ are actually new stems. The ‘branching’ is associated with those parts of the stem with creeping stolons originating from cauline apophyses mentioned above. The new stems originate from those stolons, and in all cases they begin with an intermediate internode deprived of cauline apophyses, followed by a normal series of hydrocladia-bearing internodes provided with cauline apophyses. Only in one occasion a secondary stem/branch was observed, developing from a basally and distally broken, 90-mm-high stem. Apart from a basal new stem, with intermediate internode, originating from stolons as explained above, there is a distal stem/branch originating directly from the stem through a hole in the perisarc, not from a stolon. This secondary stem differs from the new stems in being composed of a normal series of hydrocladia-bearing internodes provided with cauline apophyses (i.e. there is no basal intermediate internode).
Among the species of the genus, Oswaldella occulta sp. nov. is morphologically closer to O. tottoni in the presence of a low hydrotheca, its position on the distal half of internode, and in the number of axillary nematophores in the cauline apophyses. However, they are clearly distinguishable in several important features. The stem is divided into internodes by strongly marked nodes in Oswaldella occulta sp. nov., whereas there is no stem division in O. tottoni ; nodes are only occasionally present (cf. Peña Cantero & Vervoort 1996: 139). The degree of hydrocladial branching is also different: hydrocladia are much branched in Oswaldella occulta sp. nov., with up to six third-order hydrocladia, but only bifurcated in O. tottoni , with only two secondary hydrocladia, which, in addition, bend inwards. Another important difference concerns the separation between the prongs of the first branched hydrocladial internode and the second-order hydrocladia. In Oswaldella occulta sp. nov. there is always a distinct node between the prongs and the first internode of the secondary hydrocladia ( Fig. 7A View FIGURE 7 ), whereas in O. tottoni one, or both, of the prongs are frequently fused with the first internode of the second-order hydrocladia ( Fig. 6A View FIGURE 6 , see also Fig. 2a View FIGURE 2 in Peña Cantero & Vervoort 1996). In O. tottoni , as it was characterized by Peña Cantero & Vervoort (1996), the first hydrocladial internode, the branched one, lacks mesial inferior nematophore (cf. Fig. 6B View FIGURE 6 , see also Fig. 2a–b View FIGURE 2 in Peña Cantero & Vervoort 1996 and Fig. 12A, C in Peña Cantero et al. 1997); we can also confirm this observation, although occasionally that nematophore was present ( Fig. 6A View FIGURE 6 ). Another important difference is related to the position of the mesial inferior nematotheca, which is located at the most external point of the infrathecal swelling and its abcauline wall is distinctly directed outwards in Oswaldella occulta sp. nov. ( Fig. 7C–F View FIGURE 7 ), whereas in O. tottoni , and the remaining species of the genus with infrathecal swelling, it is located on the upper side and its abcauline wall is not directed abcaudally. Oswaldella tottoni and Oswaldella occulta sp. nov. also differ in the shape of the hydrotheca. Both species have low hydrothecae, but in O. tottoni the diameter of the hydrotheca increases from the base to the top ( Fig. 6C View FIGURE 6 ), whereas in Oswaldella occulta sp. nov. the hydrotheca has a swollen basal half and a narrower distal one, although markedly widening again at aperture, producing a sort of narrow neck with flared rim ( Fig. 7C, E View FIGURE 7 ). Oswaldella occulta sp. nov. also has distinctly thicker perisarc (compare Fig. 6C View FIGURE 6 to Fig. 7C View FIGURE 7 ), which gives it a slight stony appearance.
By the absence of ‘mamelons’ and the presence of two axillary nematophores on the cauline apophyses, Oswaldella occulta sp. nov. is also similar to O. bifurca ( Hartlaub, 1904) , O. billardi Briggs, 1938 , O. encarnae Peña Cantero, Svoboda & Vervoort, 1997 , O. erratum Peña Cantero & Vervoort, 1997 , O. incognita Peña Cantero, Svoboda & Vervoort, 1997 and O. rigida Peña Cantero, Svoboda & Vervoort, 1997 . Of these species, only O. bifurca and O. encarnae have a low hydrotheca, the remaining species have an elongate hydrotheca, being easily distinguished from O. occulta sp. nov. Oswaldella bifurca differs from O. occulta sp. nov. by the shape of the hydrotheca, which has a frontally depressed aperture, and its position on the basal half of the internode. Oswaldella encarnae clearly differs by the unbranched hydrocladia, the position of the hydrotheca on the middle of the internode, and the size and shape of the hydrotheca, which has an abcaudally directed aperture.
The material described as Oswaldella sp. by González Molinero & Peña Cantero (2015) likely belongs to O. occulta sp. nov. It is morphologically alike and was also collected off Cape Adare. González Molinero & Peña Cantero (2015) indicated that Oswaldella sp. has usually bifurcated hydrocladia with two secondary hydrocladia, although a third-order hydrocladium was not uncommon. A re-examination of the material has allowed us to put into evidence, despite the low number of hydrocladia left, that the presence of two third-order hydrocladia (one on each secondary hydrocladium) is quite common; on one occasion even two tertiary hydrocladia on the same secondary hydrocladium were observed, which indicates a likely higher degree of hydrocladial branching. The material of Owaldella sp. also shares with Oswaldella occulta sp. nov. the division of the stem into internodes, the presence of two axillary nematophores on the cauline apophyses, the position of the hydrotheca on the distal half of internode, the size and shape of the hydrotheca, and the relatively large mesial inferior nematotheca.
Distribution. Oswaldella occulta sp. nov. is currently considered endemic to the Ross Sea.
Etymology. The species name occulta comes from the Latin occultus, meaning hidden, concealed, and refers to the fact that the species was hidden among material of O. terranovae and was not noticed by Totton (1930) when he first studied this material.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |