
Ranidae Rafinesque
|
publication ID |
https://doi.org/10.11646/zootaxa.3661.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:448C4455-5A22-4C99-AA04-6FAF6DAFB879 |
|
persistent identifier |
https://treatment.plazi.org/id/B64F87EA-2064-FFE4-FF20-2A6159C8BD08 |
|
treatment provided by |
Felipe |
|
scientific name |
Ranidae Rafinesque |
| status |
|
Family: Ranidae Rafinesque View in CoL
North African Frog, Pelophylax saharicus (Boulenger 1913)
Figs. 15G–I View FIGURE 15 .
Background information. The systematic history of Pelophylax in North Africa has been subject of considerable confusion. Benhachem and Benazzou (1992) summarized relevant information concerning this topic for the Moroccan populations. Pelophylax saharicus has been suggested to be the sister taxon of either Pelophylax perezi ( López-Seoane 1885) (e.g. Uzzell 1982), or Pelophylax ridibundus ( Pallas 1771) (e.g. Pasteur & Bons 1959). Recent phylogenetic analyses have confirmed the sister relation with P. perezi (e.g. Plötner 1998; Pyron & Wiens 2011), while considerable differentiation shown by means of mitochondrial and allozyme evidence suggest that these species diverged as a result of the Gibraltar Strait opening ( Busack & Lawson 2008). The low degree of morphological differentiation between P. perezi and P. saharicus recovered by Busack and Lawson (2008) can be explained by sampling bias, with most analysed individuals originating from north-western Morocco. A similar pattern was found by Benhachem (1988), who found high similarity between P. perezi and northern P. saharicus , thus suggesting the presence of P. perezi in northern Morocco. In contrast, the occurrence of different phenotypes within Moroccan P. saharicus has been repeatedly described (e.g. Bons & Pasteur 1959; Bons & Geniez 1996), which led some authors to suggest the existence of multiple taxa. However, these phenotypes do not show any genetic differentiation ( Buckley et al. 1996; Harris & Carretero 2003), and most likely represent ecotypes shaped by different environmental factors. A similar situation has been described in Tunisian populations of P. saharicus by Amor et al. (2009).
Recent phylogenetic analyses have shown that P. saharicus occurs in entire North Africa (e.g. Arano et al. 1998; Harris & Carretero 2003). Hemmer et al. (1980) showed a wide hybrid zone between two morphologically indistinguishable ‘forms’ to be present across the Maghreb, based on electrophoretic analyses. Buckley et al. (1994) recovered similar results as the former study. This led to the designation of the subspecies P. s. riodeoroi for the western form by Arano et al. (1998), which was subsequently shown to occur in entire Morocco by Harris and Carretero (2003).
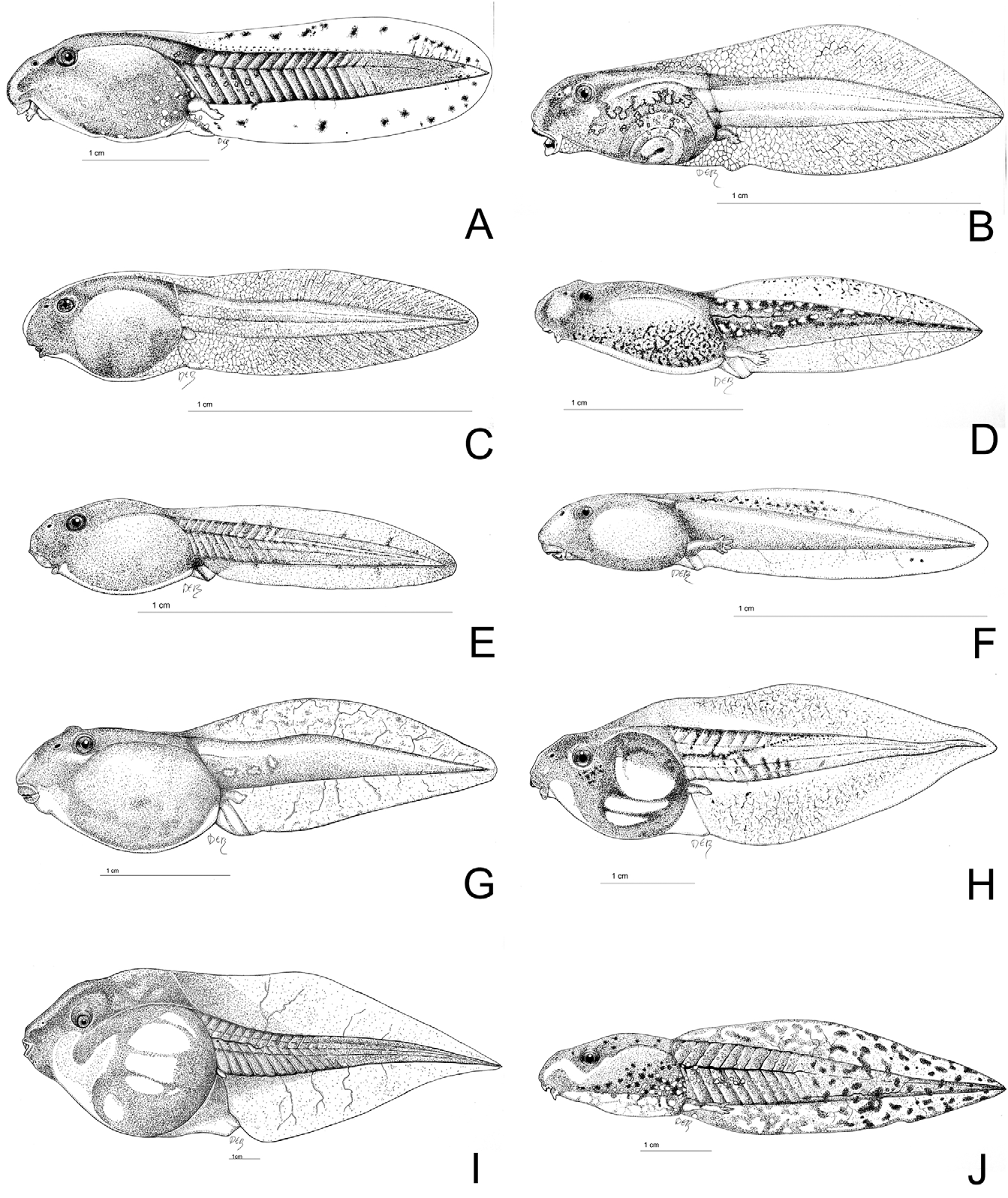
Tadpole. The tadpole of P. saharicus , based on Moroccan material, was described by Llorente et al. (1996). A drawing of the tadpole is shown in Fig. 12J View FIGURE 12 . Fully developed tadpoles are characterized by an elongated body shape and tail which is more than twice as long as the body. Eyes positioned dorsally on head. Spiracle sinistral. Oral disc ventral. Mouth almost completely surrounded by papillae, except for the upper part. One tooth row on the upper and three on the lower labium. Upper row on infralabial occasionally with narrow median gap. Colour ivory, brown or reddish with small, light brown patches on the dorsum and tail, which fade at the edges. Conspicuous single dark brown or black line runs from the anterior part of the body along the sides of the tail, to approximately two-thirds of its length.
Bioacoustics. Steinwarz and Schneider (1991) provided a comprehensive description of the advertisement call and various territorial calls of Tunisian P. saharicus . A summary of the advertisement call is presented here. Call characteristics are highly dependent of water temperature. At 19ºC, average call duration is 514.83 ms, while the call consists of 10.64 notes averaging 7.35 pulses per note. Note duration is 34.45 ms, note interval 16.62 ms. Calls can be emitted separately or in series. Although the advertisement call significantly differs from that of P. perezi in several characteristics, calls of both species are overall highly similar ( Steinwarz & Schneider 1991).
Natural history. Pelophylax saharicus can be found near mostly permanent water bodies, ranging from ponds, rivers, (mountain)brooks, oases, sources or artificial basins ( Le Berre 1989). Tolerance to human impact or alteration of habitats is particularly high. Individuals generally bask in the sun on the edge of water bodies during the day, while they venture into the surrounding terrain at night to forage ( Doumergue 1901). Insects, other frogs (including H. meridionalis ) and fishes comprise prey items ( Doumergue 1901). Bons and Bons (1959) encountered large tadpoles during the first week of April on the Doukkala plain. P. de Pous and W. Beukema developed tadpoles in a large spring within the Beni Snassen Massif in early April 2009 as well as December 2012, and near Oualidia on the Doukkala Plain in mid February 2009. Donaire-Barroso and Bogaerts (2003b) encountered tadpoles during January in the western Rif Mountains, while tadpoles were observed in the same region during September (D. Donaire-Barroso pers. obs.). Schweiger (1992) observed recently metamorphosed juveniles near Ouarzazate during late summer and autumn. These limited data suggest that at least in the northern part of the country or within the main mountain ranges reproductive activity takes place in late spring and summer, while larvae generally hibernate. Populations in the arid southern part of Morocco remain active year-round (pers. obs.).
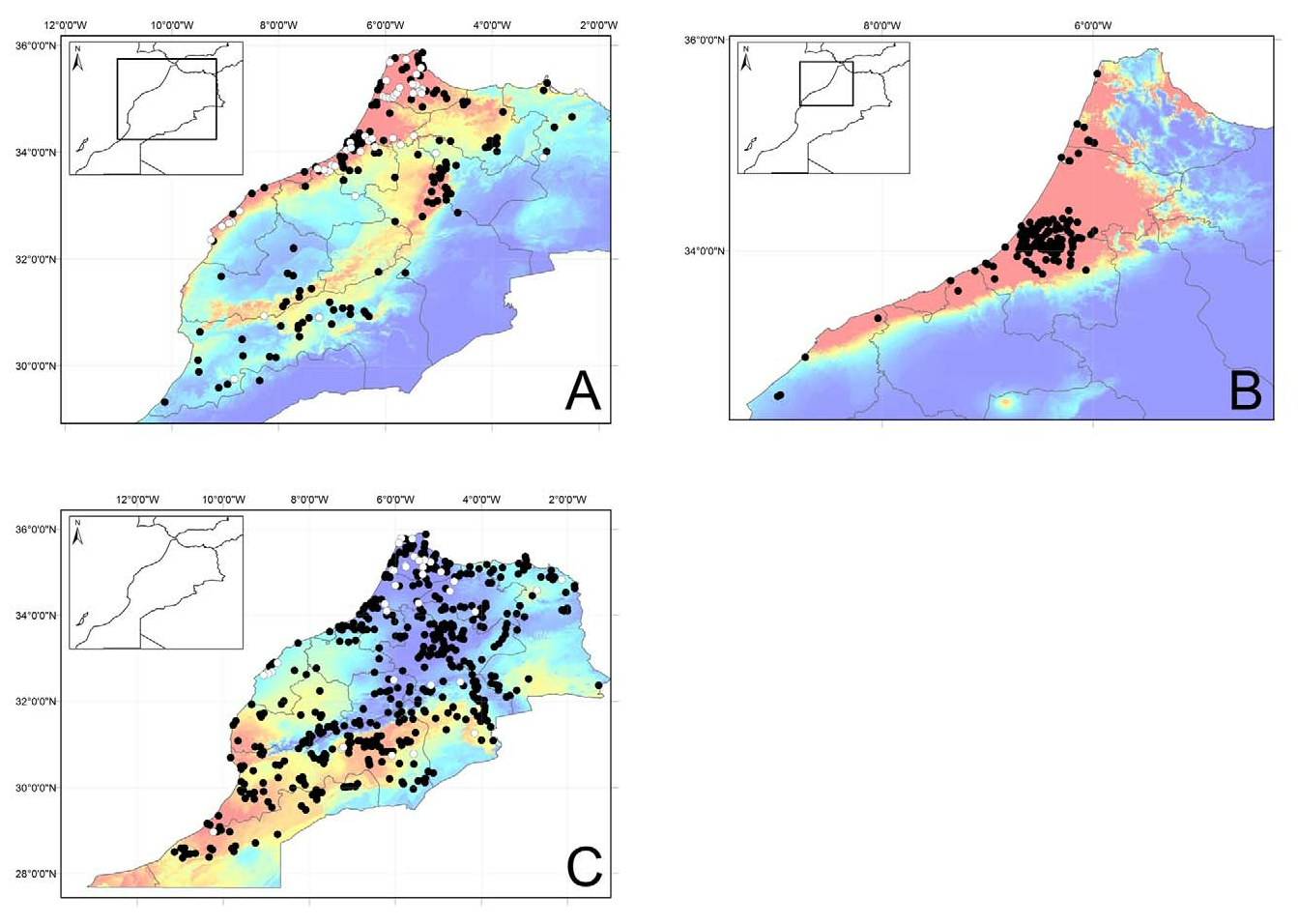
Distribution. Pelophylax saharicus is the most commonly recorded amphibian species in Morocco. The distribution map ( Fig. 16C View FIGURE 16 ) is composed of records from Bons and Geniez (1996), Brito (2003), Fahd et al. (2007), Harris et al. (2008), Ramos and Díaz-Portero (2008), Harris et al. (2010) and Barata et al. (2011). New records fill in gaps within the already vast range of P. saharicus in Morocco.
National Red List Status. Least Concern.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.