
Osteospermum
|
publication ID |
https://doi.org/10.1016/j.sajb.2022.09.001 |
|
DOI |
https://doi.org/10.5281/zenodo.10564925 |
|
persistent identifier |
https://treatment.plazi.org/id/B91BC43F-5710-E363-FF92-1349FF6D36F7 |
|
treatment provided by |
Felipe |
|
scientific name |
Osteospermum |
| status |
|
4.3. Osteospermum View in CoL
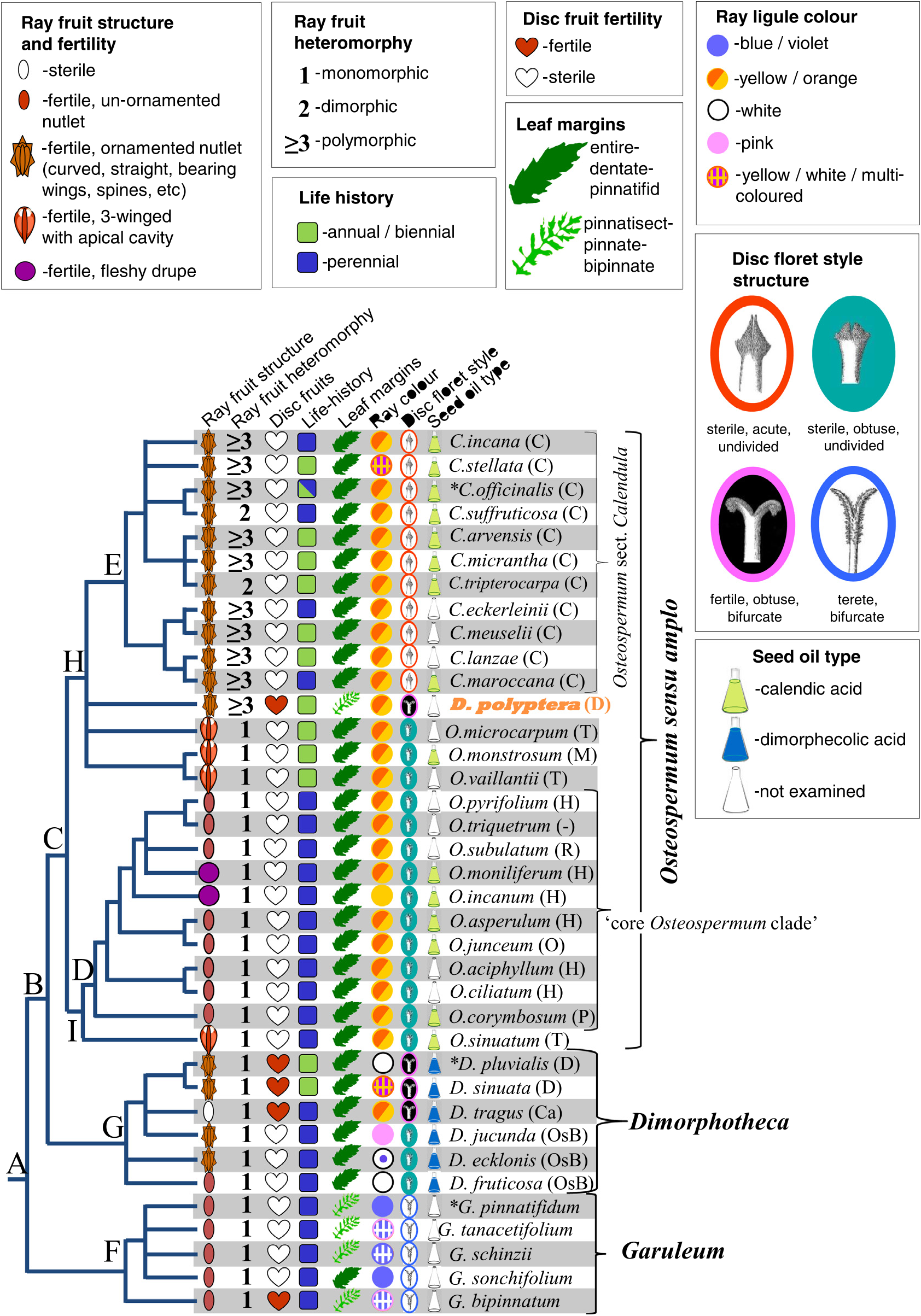
Our phylogeny is consistent with the enlarged circumscription of Osteospermum sensu ( Manning and Goldblatt, 2012) , since all sampled members of Osteospermum , Chrysanthemoides , Tripteris , and other subsequently segregated genera fall into clade C. We also demonstrate the monophyly of Osteospermum sect. Calendula based on a sample of ca. two-thirds of the species, confirming that it is embedded within clade C where it shares a common ancestor with several Osteospermum species (node H). This placement of the former genus Calendula was first demonstrated by Nordenstam (1994a) and later confirmed using DNA sequence data from both plastid (Nordenstam et al., 2006) and nuclear partitions (Nordenstam and K allersj € € o, 2009). The names Calendula and Osteospermum were published simultaneously ( Linnaeus, 1753) and thus compete for priority. Understandably, Nordenstam and colleagues were reluctant to lose the name Calendula , since members of the genus, particularly C. of fi cionalis, have a long and ongoing history of medical, culinary, pharmacological, cosmetic and religious uses in the Northern Hemisphere (e.g. https://en. wikipedia.org/wiki/ Calendula ). They were equally reluctant to lose the name Osteospermum , since synonymising this large genus would result in many new combinations. Instead, Nordenstam (1994a, b, 1996, 2006), Nordenstam et al. (2006), and Nordenstam and K€ allersj€ o (2009) attempted to maintain the monophyly of Osteospermum by dividing it into numerous smaller, monophyletic genera. The largest of these was Tripteris ( Nordenstam, 1994a) , but as indicated in Fig. 4 View Fig , the fruit type defining this genus has arisen at least twice, and possibly three times, even though we included only four of the ca. 20 species. This non-monophyly may have underpinned (Nordenstam’ s, 2006) subsequent segregation of Tripteris into the smaller genera Monoculus , Norlindhia , and Inuloides . Of these four genera, we have representatives of only Tripteris and Monoculus in our tree. The molecular phylogeny presented by Nordenstam and K€ allersj€ o (2009) resolved representatives of genera Gibbaria (2 species), Inuloides (1 species), Nephrotheca (1 species), Norlindhia (3 species), and Oligocarpus (2 species), with high support inside a clade that corresponds to our Osteospermum sensu amplo clade. However, we were unable to obtain material or sequences of species from these genera, and clearly, the phylogenetic relationships of many more species of Osteospermum sensu amplo need to be resolved.
In addition to species from the former genus Tripteris , many other Osteospermum species have fruits in which the angles are developed to a lesser or greater extent into wings, although they lack the air cavity. These species were placed by Norlindh (1943) in sections Trialatae and Spinosae of subgenus Osteospermum , the latter unfortunately with no representatives in our tree, since it contains the type species of Osteospermum , O. spinosum L.
Greater sampling, of both taxa and of genetic partitions, will be required to properly elucidate relationships within Osteospermum sensu amplo . However, phylogenetic evidence from this and from past studies indicates that the circumscription within, and relationship amongst, the clades defining Garuleum , Dimorphotheca , and Osteospermum sect. Calendula , are unlikely to change with future phylogenetic discoveries. Also, since the publication of Manning and Goldblatt (2012) resulted in the prioritisation of Osteospermum over Calendula (Turland et. al. 2018, Art. 11.5), and since all phylogenetic evidence to date indicates that Osteospermum sensu amplo is monophyletic, the enlarged circumscription of Osteospermum is likely be the most stable.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |