
Khoratichthys gibbus
|
publication ID |
https://doi.org/10.1080/02724634.2016.1225747 |
|
DOI |
https://doi.org/10.5281/zenodo.6076727 |
|
persistent identifier |
https://treatment.plazi.org/id/BA086B74-1571-FF93-FC1F-FF4EFE9431E3 |
|
treatment provided by |
Plazi |
|
scientific name |
Khoratichthys gibbus |
| status |
|
KHORATICHTHYS GIBBUS , sp. nov.
( Figs. 1 View FIGURE 1 C, 2–6)
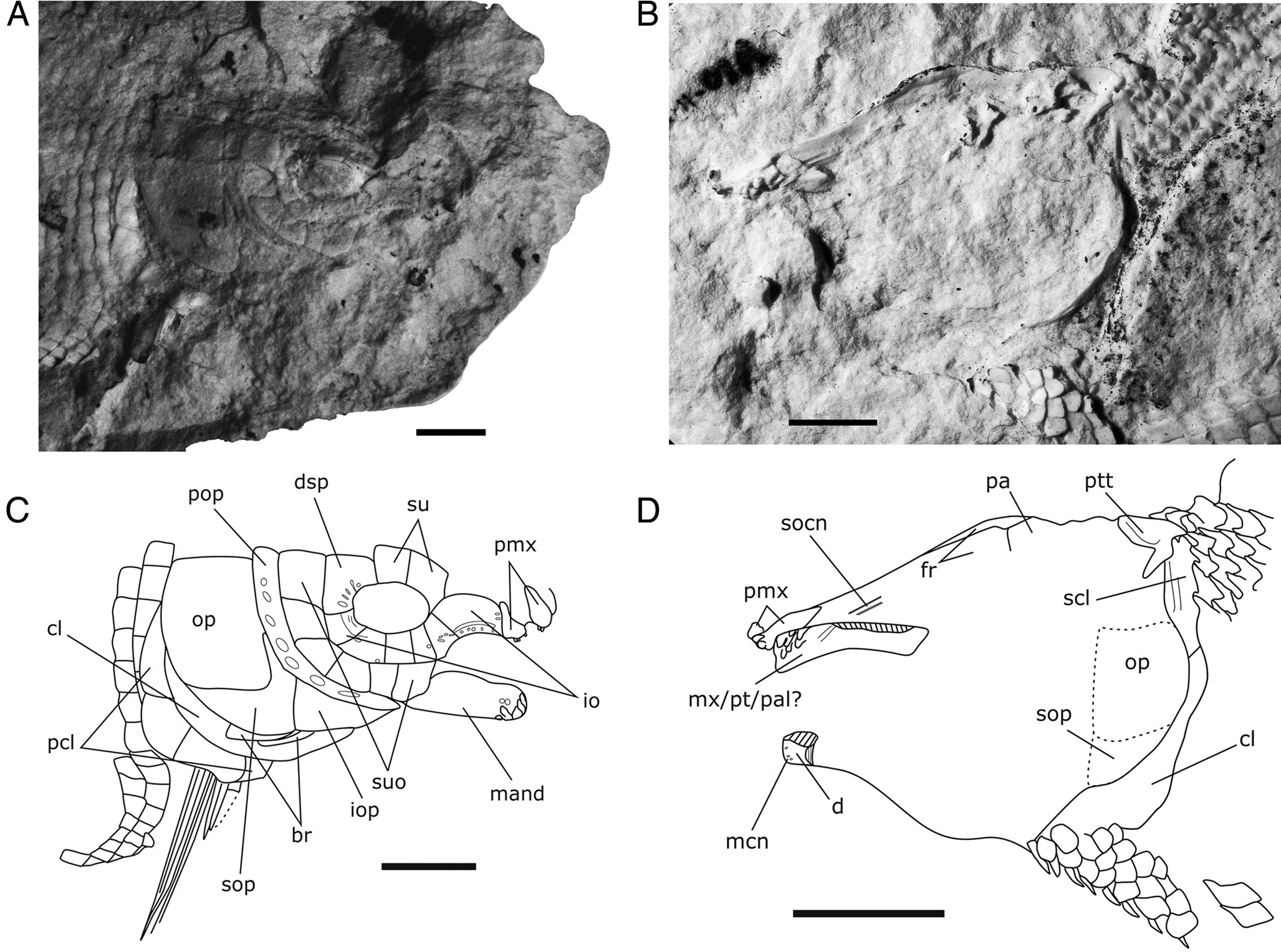
Holotype — NRRU 6011-01 , part and counter part of a subcomplete specimen preserved as negative imprints ( Fig. 1 View FIGURE 1 C).
Diagnosis — Medium-sized Ginglymodi , up to ca. 30 cm in standard length, with the following combination of morphological features: skull roof slightly concave at the level of the frontal in lateral view; nape upraised abruptly and steeply; skull bones smooth, lacking ornamentation; frontal long, frontal to parietal length ratio ca. 2.5; skull length to orbit length ratio ca. 5; closed orbital ring; few infraorbitals (six), the elongated anterior-most infraorbital with a tiny contact with the orbit; two squarish supraorbitals, the posterior one contacting the dermopterotic; large dermosphenotic; six suborbitals arranged in one row; cheek region completely covered by bones; gently curved preopercle; large interopercle; semitritorial dentition; 36 rows of ganoid scales along the lateral line and approximately 23 scales in the transverse row at the deepest level of the body; well-developed median dorsal row of scales with posterior spine; two large preanal scales; unpaired fins located posteriorly, close to caudal peduncle; narrow caudal peduncle; and ca. six pairs of long and slender fringing fulcra on the dorsal lobe of caudal fin.
Etymology — From Latin ‘ gibbus ,’ a hump.
Type Locality — Khao-Phu Luang national forest, Wang Nam Khieo, Nakhon Ratchasima, Thailand. The exact original location is unknown, but the specimen comes with certainty from the Phu Kradung Formation, probably from its upper part.
Description
The general outline of the fish shows gently curved ventral and dorsal borders, with a distinctly abrupt hump in the nape. The standard length is ca. 300 mm, and the maximum body depth, measured midway between the insertion of the pectoral and pelvic fins, is 115 mm. The head is 95 mm long including the opercular series, and 65 mm in depth. The orbit is relatively large, with a longitudinal diameter of 20 mm. The ossifications of the head have no ornamentation except on the first and the second infraorbitals, which bear some tiny tubercles restricted to the ventral border of the bones. Both dorsal and anal fins are located close to caudal peduncle (predorsal ca. 200 mm, preanal ca. 225 mm). The level of insertion of the dorsal fin is situated just in front of the level of insertion of the anal fin. The dorsal ridge scales are large and bear elongate posterior spines. The caudal peduncle is relatively narrow compared with the body depth, approximately 3 times shallower (caudal peduncle ca. 40 mm).
Skull Roof — The frontal, parietal, extrascapular, and dermopterotic are not directly visible. However, the trace on the counterpart of the skull shows the general outline of the frontal and parietal.
The length of the frontal, which is slightly concave in its anterior part, is approximately 45 mm. The surface of the bone, as observed from the imprint, shows no trace of ornamentation. Although not completely visible, the frontal appears to be proportionally long, probably 4 times longer than wide. The parietal measures ca. 15 mm in length. Its surface is smooth.
Details of the extrascapulars are difficult to see, and our reconstruction of this bone is tentative. At least one extrascapular can be identified as a gently curved groove on the counterpart corresponding to the anterior limit of the bone. The nasal bone is not visible.
Circumorbital and Suborbital Series — The circumorbital series is composed of a large dermosphenotic, two supraorbitals, and at least six infraorbitals located ventral and anterior to the orbit. The first, anterior-most infraorbital is the largest of the series. Although this ossification has a tiny contact of its posterodorsal corner with the orbit ( Fig. 3 View FIGURE 3 A, C), we consider that this infraorbital is homologous to the anterior infraorbital(s) without contact with the orbit of other ginglymodians, which is a synapomorphic character of the clade ( López-Arbarello, 2012). This first infraorbital is longitudinally elongate, roughly rectangular in shape, about 1.7 times longer than deep, with dorsal and ventral margins gently curved. The surface of this bone is smooth but close to its ventral margin there is a narrow curved groove. Under that curved groove are several regularly spaced rugae running along the ventral edge of the bone. The second infraorbital is approximately trapezoidal in shape and is deeper than long, with its ventral margin longer than the dorsal margin. Regularly spaced rugae occur in continuation with those from the first infraorbital, but we cannot observe the continuation of the groove. The third rectangular infraorbital is the smallest bone of the circumorbital series. The three remaining infraorbitals are approximately square in shape, with their dorsal and ventral margins apparently curved. Their surfaces are smooth, but the dorsal-most infraorbital presents a ridge running along its dorsal margin that reaches the dermosphenotic. The shape of the dermosphenotic is approximately trapezoidal, with its anteroventral margin concave, following the rim of the orbit. Close to the anteroventral margin of the bone at least three nodes are present arranged in the continuation of the ridge on the posterior-most infraorbital, indicating the path of the infraorbital sensory canal and its connection with the otic sensory canal. Two supraorbitals are situated dorsal to the orbit. The anterior-most is the largest.
It is square and contacts the anterior-most infraorbital. The posterior one is approximately rectangular and deeper than long. It seems that both supraorbitals bear tiny tubercles restricted to their ventral edges and mostly visible on the anterior one. There are six suborbitals arranged in a single row. The surface of these bones lacks ornamentation. Their shapes are rectangular but they show variations; the anterior-most one is subrectangular, with its ventral margin forming a curved line, whereas the other margins are straight. The second suborbital is approximately rectangular, deeper than long, with its anterior margin longer than the posterior margin. The third suborbital is elongated, rectangular, with a gently concave dorsal border and a convex ventral border. The fourth and fifth suborbitals are quite similar in shape, i.e., subrectangular, with the dorsal border shorter than the ventral border and the ventral margins gently convex. The dorsal-most suborbital is approximately square and is the largest of the series.
Jaws — The dentary is partially preserved. Only the anterior portion of the bone with a pack of teeth is visible ( Fig. 3 View FIGURE 3 A–D). The visible part shows parallel curved margins. The teeth on the dentary ossification are apparently arranged in four lines, with possibly the internal rows borne by the coronoid bone. Each tooth is composed of a cylindrical base, 1 mm high, and of a small bulbous tiny acrodine apex ca. 2 mm high. A bone of the upper jaw with seven teeth is visible on the counterpart of the silicone peel. This thin and elongate bone is possibly the maxilla. The dentigerous part is restricted to the anterior portion. An alternative interpretation of this ossification is that it corresponds to a compound structure including part of the pterygoid and palatine. The premaxillary ossification is relatively small, with its anterior rim rounded. Posteriorly, the narrow and elongate nasal process of the premaxilla seems to extend under the frontal. There are several teeth preserved on the premaxilla, but their exact number is unknown. The teeth are similar in shape, but slightly smaller, than the teeth borne on the dentary and maxilla. Based on the location of the lower and upper jaws, it seems that this fish had a wide gape.
Opercular Series — The opercular series is complete, formed by the preopercle, opercle, subopercle, and interopercle ( Fig. 3 View FIGURE 3 A–D). The preopercle is a distinctly narrow and curved bone. The bone seems to have parallel lateral margins, except at the posteroventral corner, which shows a gently convex contour, making the mid-depth of the bone slightly broader. There is no tapering dorsally, and the ossification ends anteriorly as a blunt spine. A series of seven large pores located in the center of the bone represent the preopercular sensory canal. The opercle is approximately square in shape, about 1.2 times deeper than long (32 X 25 mm). Its anterior border is straight, as is the dorsal one, whereas the posterior border is gently convex. The external surface is smooth. Ventrally, the opercle fits with the dorsal margin of the subopercle. The subopercle is relatively shallow, with its maximum depth about 0.4 of the depth of the opercle. The anterior ascending process of the subopercle is relatively low; its height is less than half depth of the opercle. Posterior to the ascending process, the subopercle narrows posterodorsally, so that the dorsal border of the bone is concave whereas the ventral border is convex. Anteriorly, the subopercle articulates with the interopercle through a vertical contact. The interopercle is visible as a large triangular bone; its maximum depth and length are equal to those of the subopercle. Anteriorly, it extends up to the level of the anterior extremity of the preopercle.
Hyoid Arch and Branchiostegal Rays — We cannot distinguish the ceratohyals. Similarly, the number of branchiostegal rays is difficult to establish, but at least two can be observed lying under the subopercle.
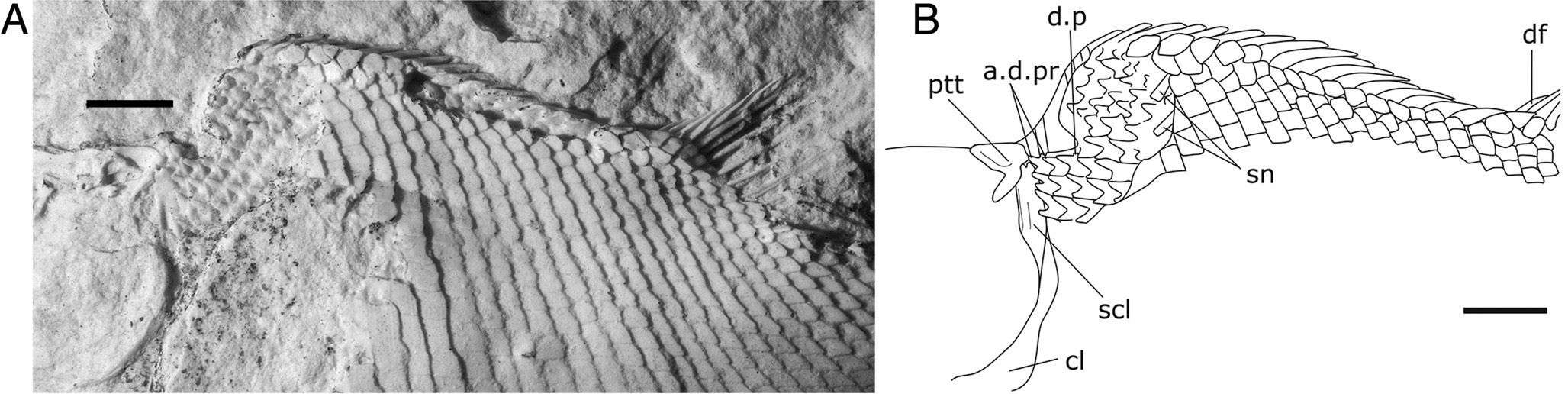
Pectoral Girdle — The posttemporal and the supracleithrum are visible on the counterpart from the internal side only ( Fig. 3 View FIGURE 3 B, D). The posttemporal appears as a deep, crescentshaped bone excavated anteriorly. The supracleithrum, only partially visible, shows a vertical groove situated close to the anterior margin of the bone, which likely corresponds to the path of the supratemporal sensory canal. The right cleithrum and the postcleithrum are visible, but the limits of the bones and especially the anterior portion of the cleithrum are not clearly seen. There are four postcleithra, all with a smooth surface. The anteroventral-most one is the smallest of the series ( Fig. 3 View FIGURE 3 A, C).
Cephalic Sensory Canal — The supraorbital sensory canal is visible as a groove on the internal side of the frontal on the silicone copy of the left side ( Fig. 3 View FIGURE 3 B, D). Because only the anterior trace of this groove is visible, it is uncertain if this canal runs through the parietal or the dermopterotic. Only the paths of the infraorbital sensory canal and of the preopercular sensory canal are visible on the right side of the specimen. There is no evidence of a sensory canal running within the supraorbital bones. The preopercular sensory canal is present as a series of large pores located in the center of the ossification ( Fig. 3 View FIGURE 3 A, C). The mandibular sensory canal can be recognized as at least three pores arranged in one line close to the dorsal margin of the dentary ( Fig. 3 View FIGURE 3 B, D).
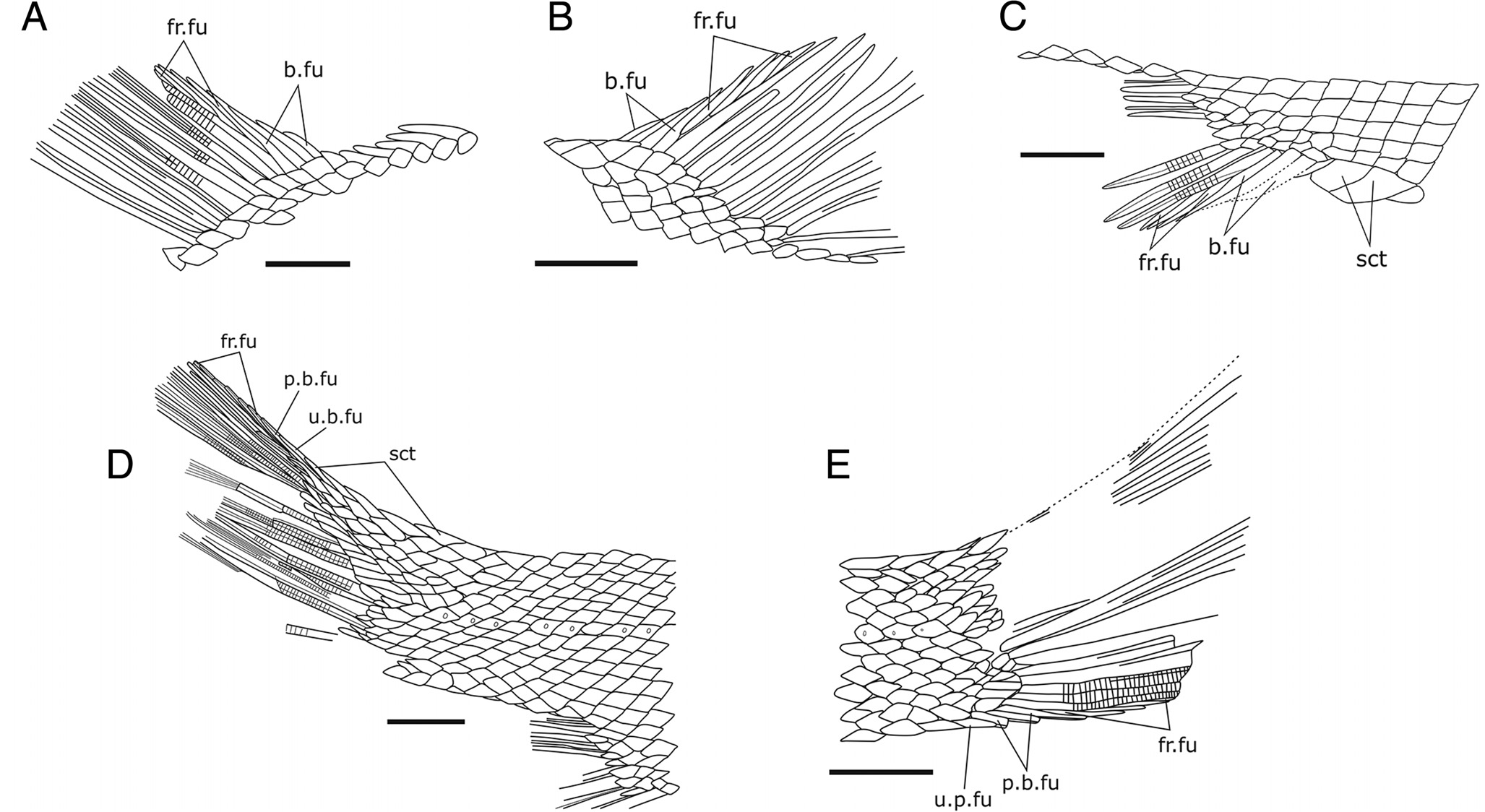
Pectoral Fins — The pectoral fin is slightly twisted anteriorly; it consists of at least three basal fulcra, the anterior one being possibly unpaired and two posterior ones being paired. Because sediment partially covers the pectoral fin, it is unclear if there are fringing fulcra. The visible five rays are very long, representing 5 2% of the head length (pectoral fin length = 5 0 mm, head length = 9 5 mm), and they appear to be unsegmented ( Fig. 3 View FIGURE 3 A, C).
Pelvic Fins — The pelvic fins are poorly preserved. They are located at the level of the ninth and 10th transverse scale rows, halfway between the pectoral and anal fins. The pelvic fin originates at the level of the fourth vertical scale rows up from the ventral midline scales ( Fig. 6 View FIGURE 6 ).
Unpaired Fins — The dorsal and anal fins are almost complete. The dorsal fin inserts a little in front of the anal fins. It is composed of four basal fulcra, approximately five fringing fulcra, and nine rays ( Fig. 4 View FIGURE 4 A, B). The proximal unsegmented portion is long; it is nearly half the fin length of the anterior ray and gradually decreases posteriorly. The anterior first ray is unbranched, the second ray branches at least once, and the following rays branch twice, with four terminal branchlets showing fine segmentation. The length of the dorsal fin is nearly half the head length. Although we cannot see the tips of the rays, we assume that the distal margin of the fin was straight.
The anal fin originates right behind the level of the 20th transverse scale rows. It is slightly smaller than the dorsal fin, comprising three basal fulcra, at least three stout fringing fulcra, and approximately 11 fin rays ( Fig. 4 View FIGURE 4 C). The proximal unsegmented portion represents one-third of the length of the rays and progressively decreases in size posteriorly. There are two large preanal scutes, which are about twice as large as the adjacent scales.
The caudal fin is also relatively complete ( Fig. 4 View FIGURE 4 D, E). There are 18 rays; eight in the ventral lobe and 10 in the dorsal lobe. The dorsal lobe is preceded by five scutes covering the body lobe of the tail, followed by two thin and elongate basal fulcra (the anterior one is unpaired), and possibly six pairs of very thin fringing fulcra lying on the dorsal marginal ray. The caudal fin rays are intensively branched, with the exception of the two marginal rays. The proximal unsegmented portion is short. Ventrally, the caudal fin is preceded by one short unpaired basal fulcrum, three paired basal fulcra, and at least four thin and small fringing fulcra.
Squamation — The body is covered with rhomboid scales, which have smooth surfaces and smooth borders. The anterior margin of each abdominal scale presents a ‘double-peg’ articulation ( Fig. 5 View FIGURE 5 A, B), with the anterior dorsal process more protruding than the anterior ventral process (visible on the postcranial region of the counterpart). In addition, there is a small process protruding from the dorsal border of the scale, which fits in a conical socket excavated in the ventral median border of the scale lying above. This arrangement indicates the presence of a vertical peg and socket articulation (visible nearly in the center of the trunk of the right side). There are 36 transverse scale rows carrying the lateral line. There are 23 vertical rows of scales dorsoventrally and 14 inverted row scales forming the body lobe of the tail. As usual in ginglymodians, the shape of the scales varies in different parts of the body, from rectangular, somewhat deeper than long in the anterior and central region of the flank, to rhomboidal in the caudal peduncle. The dorsal ridged scales are conspicuous, very large, and bear a stout posterior spine inclined posteriorly. There are approximately 18 dorsal ridged scales. Ventrally, scales from the median midline also bear a short posterior spine directed backward. The lateral line scales are characterized by a small pore on their surface. There are two stout bones exposed just behind the cranium. These two bones are referred to supraneural ossifications ( Fig. 5 View FIGURE 5 A, B). No ossified vertebral centra are visible.
DISCUSSION
The specimen from Wang Nam Khieo shows a combination of characters, listed in its diagnosis above, that justifies its inclusion in a new genus and species, Khoratichthys gibbus . It can be confidently included in the Ginglymodi on the basis of the occurrence of one synapomorphy, ‘one infraorbital anterior to the circumorbital ring (not including the antorbital),’ and of a combination of homoplasies ( López-Arbarello, 2012; Deesri et al., 2014). Other ginglymodians have a humpbacked profile reminiscent of Khoratichthys , such as the Triassic Kyphosichthys grandi from China (Xu and Wu, 2012) and the Triassic Lophionotus sanjuanensis from the United States ( Gibson, 2013b). However, K. gibbus clearly differs from K. grandi by the cheekbone pattern, the ornamentation of the dermal bone, and the morphology of the scales, among other characters. Khoratichthys gibbus also differs from L. sanjuanensis by the number and arrangement of suborbitals and infraorbitals, and by the presence of an elongated posterior spine on the dorsal ridge scales, among other characters.
Cladistic Analysis — In order to assess the phylogenetic position of Khoratichthys gibbus among the Ginglymodi , we performed a cladistic analysis of a set of ginglymodian taxa. Our aim here is not to address the ‘gar- Amia -teleost’ problem (cf. Grande, 2010, for discussion); consequently, we considered Amia calva as an outgroup for the analysis, together with Polypterus ornatus and Leptolepis coryphaenoides . Because speciesrank taxonomic issues are still pending for several ginglymodian taxa, we restricted the analysis to the type species of 25 genera. We added Isanichthys lertboosi (alongside the type species I. palustris ) because we also address here the taxic diversity of ginglymodians in the Phu Kradung Formation at the species level. The characters used in the analysis form a new set of 103 characters built on a critical combination of the characters defined by Cavin (2010), Grande (2010), López-Arbarello (2012), and Deesri et al. (2014). The character definitions, associated with discussion for some of them, are available in Supplementary Data 1, and the data matrix is available in Supplementary Data 2.
The parsimony analysis was run in PAUP* 4.0b10 ( Swofford, 2001). A heuristic search (using random addition sequence, 10,000 replications, one tree held at each iteration, and tree bisection and reconnection branch swapping) was carried out. Figure 7 View FIGURE 7 shows the strict consensus tree of the five most parsimonious trees produced (92 characters informative; 352 steps; consistency index = 0.4063; retention index = 0.6151). The list of character changes for one of the most parsimonious trees is available in Supplementary Data 3.
In the consensus tree, Ginglymodi is a well-supported node (Bremer index [BI] of 2 and bootstrap support [BS] of 58) with 12 character changes, five of them uniquely derived: the junction of supraorbital canal with infraorbital canal not in the frontal (char. 18) (but this character is coded for 35% of the ingroup taxa only), the absence of the Mentomeckelian bone (char. 72), the absence of gular bone (char. 75), the presence of a medial wing on cleithrum (char. 86), and the presence of fringing fulcra (char. 94, state 1). Only two of these characters (char. 75 and 94) are unambiguous with the ACCTRAN optimization. A premaxilla immovably attached to the braincase by means of a long nasal process tightly sutured to the frontals (char. 1) also supports the Ginglymodi node, but this character is also present in the Amia branch, a clade used as an outgroup here. The main differences between our topology and those of López-Arbarello (2012) and Gibson (2013b) are: (1) the Semionotiformes are not monophyletic, (2) the Macrosemiidae are not nested within the Semionotiformes but are the sister group of all other Ginglymodi , and (3) the Callipurbeckiidae are monophyletic in our analysis, as in the López-Arbarello (2012) and Gibson (2013b) analyses, but this family also includes Sangiorgioichthys here, whereas the latter is resolved in a more basal position in López- Arbarello’s study (2012). The unique derived unambiguous character that supports the Callipurbeckiidae is ‘two suborbitals, the ventral one much larger than the dorsal one’ (char. 42, state 4). Most of the other characters are homoplastic and not in common with López-Arbarello’s study (2012), because of different definitions of characters and/or differences in the optimization. The only exception is the occurrence of a moderately tritorial dentition (char. 56), which is homoplastic but common to both studies. The sister group of the Callipurbeckiidae in our tree comprises a polytomy including Khoratichthys gibbus , Neosemionotus, Lophionotus , and all the more derived Ginglymodi . This wellsupported node (BI of 2) is defined by several homoplasies and two uniquely derived apomorphies: quadrate/metapterygoid contact or close association (char. 47) and symplectic/quadrate articulation (char. 54), but none of these are coded in Khoratichthys gibbus , Neosemionotus , and Lophionotus and consequently both are ambiguous. The fourth clade of the polytomy, comprising Lepidotes gigas and the more derived Ginglymodi , is rather well supported (BI of 2), although all the six character changes supporting the node are homoplastic. The sister group of L. gigas comprises a trichotomy with both species of Isanichthys and the more derived Ginglymodi . The coding of characters for the two Isanichthys species differs for six characters only, two dealing with head proportions (char. 8 and 13), two with mandibular (char. 64) and body (char. 89) proportions, one with the infraorbital pattern (char. 32), and one with the occurrence versus absence of teeth (char. 61). Deesri et al. (2014) provided a discussion of the polymorphism observed in I. lertboosi and its differences with I. palustris . Further studies based on new material referable to Isanichthys are necessary for providing a better definition of this genus and its species content. The phylogenetic relationships of the more derived ginglymodians are as follows: Scheenstia ( Thaiichthys (( Araripelepidotes Pliodetes ) ( Obaichthyidae Lepisosteidae ))). This pattern is similar to the one found in Deesri et al. (2014), except that in the latter Thaiichthys is resolved as the sister group of ( Araripelepidotes Pliodetes ).
Taxonomy — Because López-Arbarello (2012) not only provided stem-based definitions of the clades but also provided character-based diagnoses, we have difficulties to name some of the clades in our phylogeny. This is especially true for the Lepisosteiformes . Thanks to the stem-based definition, Lepisosteiformes is the clade including all taxa more closely related to Lepisosteus than to Semionotus, Macrosemius, Dapedium , or Amia . In our cladogram, this definition implies that Lepisosteiformes gathers all taxa more derived than Semionotus , including the Callipurbeckiidae . Based on this definition, the composition of the lepisosteiforms is very different from López-Arbarello’s original definition of the clade. If we use López-Arbarello’s character-based diagnoses by mapping her diagnostic characters of Lepisosteiformes on our cladogram, seven characters are spread between the node supporting the Callipurbeckiidae plus more derived Ginglymodi and the node supporting Thaiichthys plus more derived Ginglymodi . The spread of these characters makes a diagnostic-based definition difficult. As a consequence, with the will to keep a conservative definition of the Lepisosteiformes (i.e., without the inclusion of the Callipurbeckiidae ), we use here a stem-based definition but amend it as follows: Lepisosteiformes is the clade including all taxa more closely related to Lepisosteus than to Semionotus, Callipurbeckia, Macrosemius, Dapedium , or Amia . Based on this definition, Khoratichthys gibbus is amongst the basal-most Lepisosteiformes , together with Neosemionotus and Lophionotus.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |