
Maabella stomalata Hastriter and Bush, 2006
|
publication ID |
https://doi.org/10.11646/zootaxa.1636.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FF92CF8E-6B12-4E10-BC65-8D7B9CA908A8 |
|
persistent identifier |
https://treatment.plazi.org/id/BE5FF613-0717-2015-FF28-F8E259D60CE4 |
|
treatment provided by |
Felipe |
|
scientific name |
Maabella stomalata Hastriter and Bush, 2006 |
| status |
|
Maabella stomalata Hastriter and Bush, 2006 View in CoL
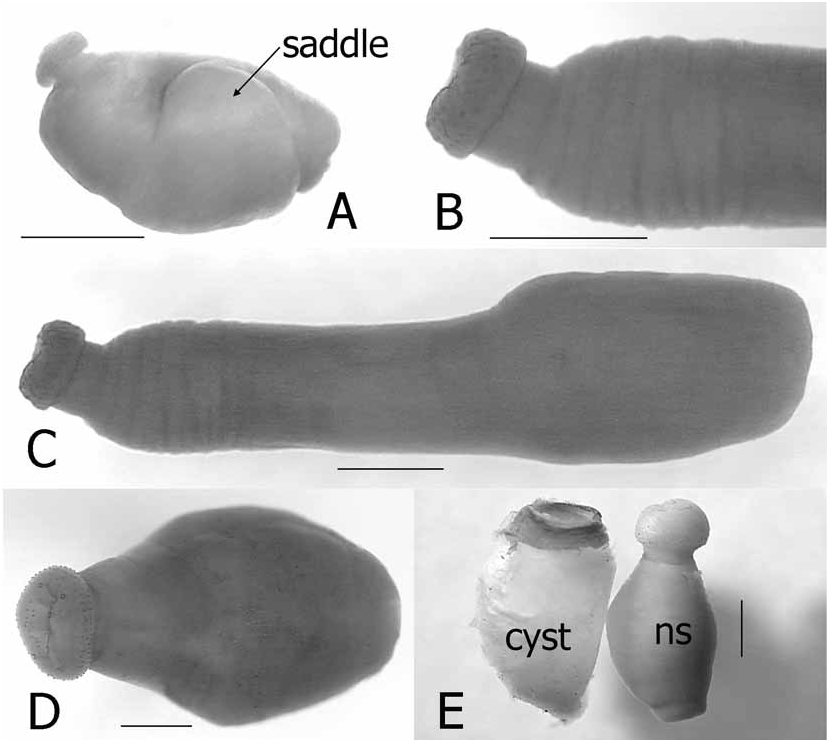
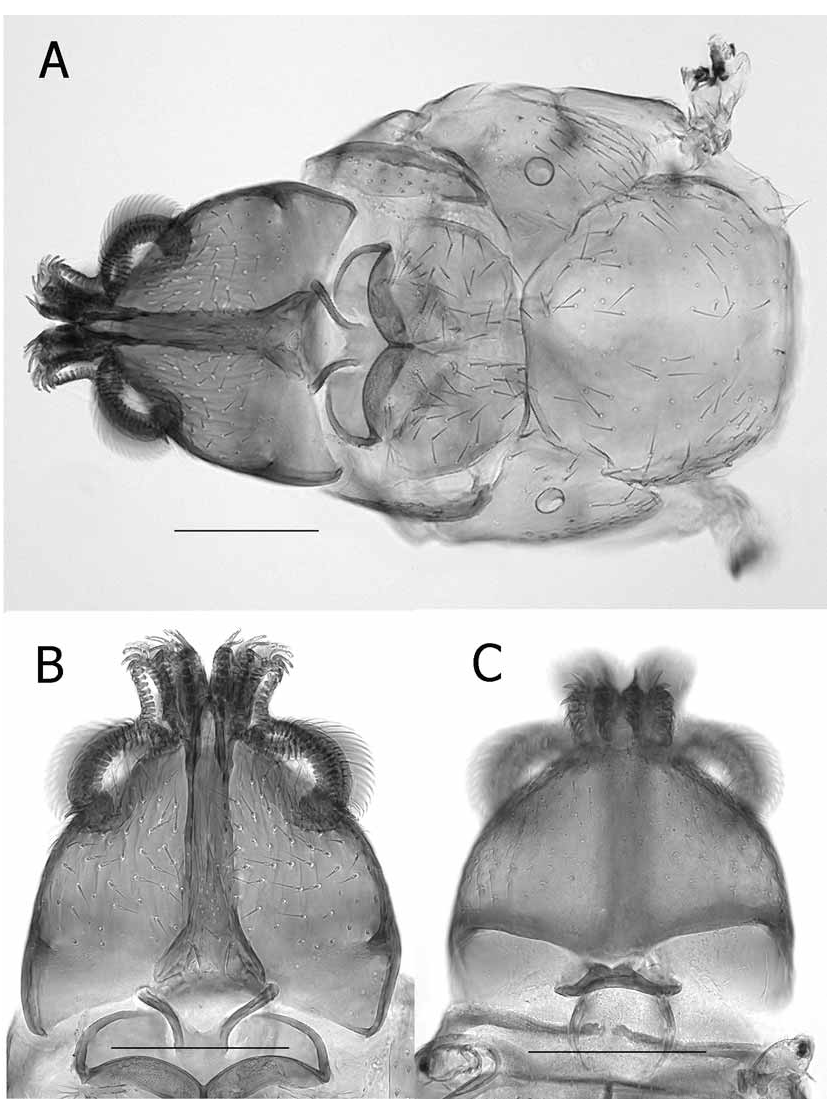
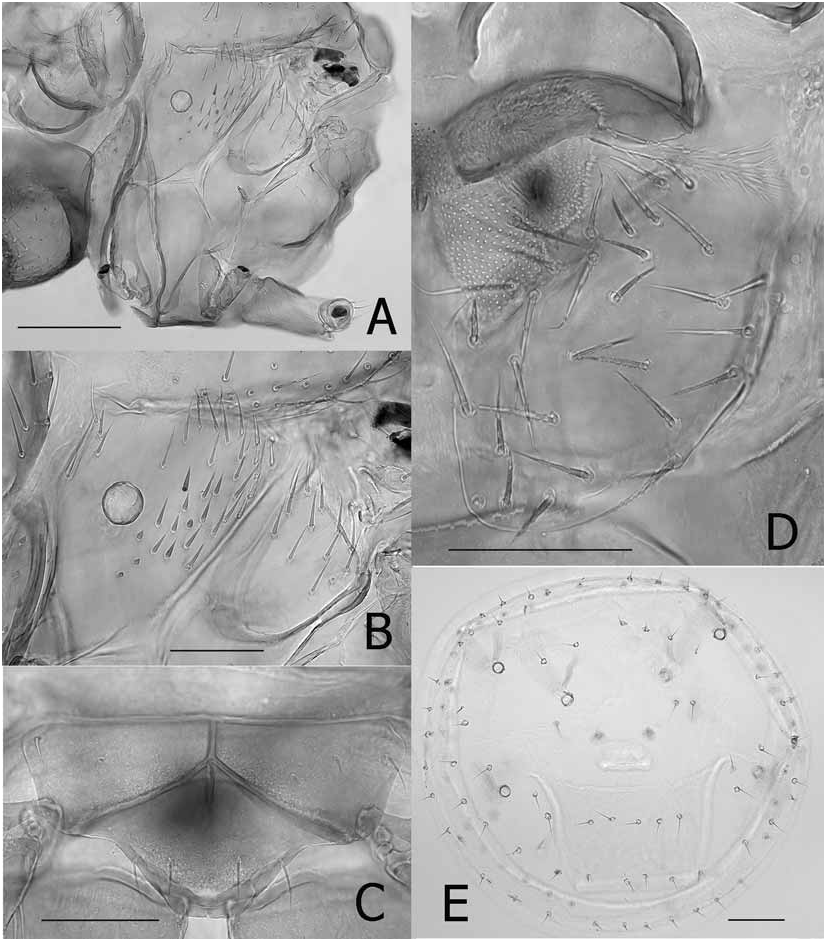
(See Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 in Hastriter and Bush, 2006)
Material examined. Specimens listed in Hastriter and Bush 2006 are not presented here. The following new records greatly expand the heretofore known range of this species. Borneo: Ex. Rhinolophus borneensis
Peters (3, BM-1007A–C, BPBM). China: Jin Xin County Provincial Nature Reserve, ( 23°07'21"N, 105°57'49"E) [Note: the coordinates for specimens from this locality were recorded erroneously in Hastriter and Bush (2006: 33) as 23°12'N, 105°09'E], 978 m, Guangxi Province, R. affinis ( V) ( F47252 View Materials ), 20 IX 2004, S.E. Bush, (1, ROM- 116026/ F47252 View Materials , ROM). Indonesia: Java: Soerabaya, ex. Rhinolophus ? philippinensis Waterhouse, (4, wing, Leiden-4000A–D, RMNH); Tjibodas, ex. Rhinolophus sp. (“possibly affinis ”), 1890, J.F. Bemmelen, (2, wing, Leiden-4001A–B, RMNH); Modjokerto (eastern Java), ex. Rhinolophus sp. , (3, wing, Leiden-4002A–B, Leiden-4003, RMNH); Solo, ex. Rhinolophus acuminatus Peters , (2, wing, Leiden- 4004A–B, RMNH); Moluccas: Amboina, ex. Rhinolophus euryotis Temminck ( V)[ RMNH #165, (Rijksmuseum van Natuurlijke Histoire), Leiden, Netherlands], (2, wing, Leiden-4007A–B, RMNH); Grand Keys, ex. R. euryotis ( V), 4 VIII 1865, H. von Rosenberg, (4, wing, Leiden-4006A–D, RMNH); Solor Island (Lessor Sunda Islands), ex. Rhinolophus rufus Eydoux and Gervais ( V) ( RMNH #167), 1864, J. Semmelink, (3, wing, Leiden-4008A–C, RMNH); Obi: Anggai, ex. Rhinolophus sp. , 23 III 1953, A.M. R. Wagner (3, 3 larvae in genitalia vials, 6 neosomes in alcohol, wing, Bogar-3001A–C, BPBM). Malaysia: Gua Anaktakun, ex. R. affinis , 26 VIII 1966, T.C. Maa, (3, wing, BM-1008A–D, BPBM); Kedah, Kodiang area, ex. R. affinis (hosts RH-131–135), 26 II 1968, (2, penis, ROM- 6000A–B, ROM); Pahang, Gua Che Vatim Cave no’s 1, 2 & 3, ex. Hipposideros sp. , 26 XII 1966, T.C. Maa and J.L. Gressitt, (3, 5 neosomes in alcohol, wing, BM-1048A–C, BPBM); Pahang, 24 km E Jerantut Ferry, ex. R. affinis , 11 IV 1967, A.J. Beck, (8, A-114A–H, BOHART). Myanmar: Karen Hills, ex. R. affinis, II 1896 , L. Fea (3, 1 larva in genitalia vial, wing, Genova-2001A–C, BPBM); Karen Hills, Taho, ex. R. affinis macrurus Andersen = R. affinis ( V) (Leiden #1567), II 1888, L. Fea (1, wing, Leiden-4005, RMNH); Cave, 9.7 km W Haibum, ex. R. affinis macrurus = R. affinis ( V) ( AMNH #112839), 5 III 1935, H.C. Raven (6, ca. 50 neosomes in alcohol, wing, AMNH-5000A–B, AMNH-5001 A-D, AMNH); Papua New Guinea: Iamai Cave, 0.6 km N Maneaoa, 16 km E Biniguni, ex. H. cervinus ( V) (A.E. #12790, #13739, 6 VIII 1953, H.M. Van Deusen (2, wing, BM-1014A–B, BPBM); Iamai and Gwebmantoi Caves, 0.6 km N Maneaoa, 16 km E Biniguni, ex. Hipposideros cervinus cervinus = H. cervinus ( V) (A.E. #12812–12879), 6 VIII 1953, H.M. Van Deusen (14, 2 larvae in genitalia vials, and ca. 220 neosomes in alcohol, wing, BM-1015A–C, BM-1016A–C, BM-1017A–B, BM-1051A–C, BM-1052A–C, BPBM. 10 neosomes in alcohol, FMNH); Louisiade Archipelago: Ehaus Caves, Misima Island, Narian, ex. Aselliscus tricuspidatus (Temminck) ( V) ( AMNH #14219), 6 VIII 1956, R.F. Peterson (1, DNA voucher) (BM-1002, BYU); same data except ex. Rhinolophus megaphyllus Gray ( V) (A.E. #14220, 14225–14227) (3, 2 larvae in genitalia vials, 3 neosomes in alcohol, wing, AMNH-5002A–C, AMNH); Nissan Island (Green Islands), ex. Hipposideros galeritus Cantor ( V) ( AMNH #99907), VIII 1927, H.D. Hamlin (Whitney Expedition) (3, 1 larva in genitalia vial, AMNH-5004A–C, AMNH). Philippine Islands: 16 km SE San Nicolas, Busuanga Island, ex. “leaf-nosed bat”, 22 V 1962, Max Thompson, (5, BM –1005, BM-1006A–B, BM-1018, BBM- 2982, BPBM); 6 km NE San Nicolas, Busuanga Island, ex. Rousettus amplexicaudatus (E. Geoffroy) ( V) ( BBM-PI #3011), 24 V 1962, Max Thompson (3, 1 neosome in alcohol, BM-1027A–C, BPBM); same except ex. Rhinolophus sp. ( V), 26 V 1062, (1, BM-1031, BPBM); Basay Limestone caves, Negros Oriental Province, Negros Island, ex. Hipposideros sp. , 12 XII 1959, L.W. Quate, (3, 2 immature larvae, 1 discarded, 1 in genitalia vial, 3 neosomes in alcohol, wing, BM-1039A–C, BPBM); Cebu Minglanilla, SE Ceba City, 500- 600m, limestone cave, ex. Hipposideros sp. , 4, 5 II 1962, L. Quate and S. Quate, (6, 5 neosomes in alcohol, wing, BM-NG-891–895A–C, BM-NG-896, BM-NG-901–903A–B, BPBM). West Papua: Humboldt Bay, Grotto, nr Lake Santani, ex. H. calcuratus (Dobson) , 9 IV 1903, Humboldt Bay Expedition, (3, 1 mounted larva, 13 neosomes in alcohol, wing, Leiden-4010A–C, RMNH); Cave at Djidmaoe, Vogelkop, ex. Hipposideros sp. , 2 VI 1955, A. Boeseman, (3, 1 mounted larva, 17 neosomes in alcohol, wing, Leiden-4013A–C, RMNH); Hollandia, ex. Hipposideros cervinus cervinus = H. cervinus ( V) ( AMNH #152705–152708), 6 VII 1938, Archbold Richardson (3, wing, AMNH-5003 A-C, AMNH); Djidmaoe, Vogelkop, ex. Hipposideros sp. , 13 VI 1952, Aangebrecht Door Bevolking Expedition, (1, wing, BM-1030, BPBM); “West Papua or Papua New Guinea, mixed data”, ex. Hipposideros sp. , (1, wing, BM-1041, BPBM); Ajappo (Eastern peninsula of Lake Santani), Hollandia, ex. Hipposideros sp. , 6 XI 1954 (3, 2 immature larvae in genitalia vials, wing, BM- 1043A–C, BPBM); Wersar, ex. Hipposideros sp. ), Mrs. Marcus v.d. Nieuwenhuiser (3, 1 mature larva mounted, 2 immature larvae discarded, wing, BM-1047A–C, BPBM); Ajappo (cave), south side of Lake Santani, ex. Hipposideros sp. , (3, larva in genitalia vial, wing, BM-NG-568–582A–C, BPBM).
Diagnosis and description. see Hastriter and Bush (2006).
Dimensions. These are provided in Hastriter and Bush (2006) except for those of the neosome and genital aperture, which are provided herein: Length of neosome: 2500µ (n = 53, range: 2000–4000µ). Genital aperture, diameter: 632µ (n = 14, range: 464–803µ) ( Table 1).
Remarks. The description of M. stomalata was based on material limited to China and Vietnam. In this study, additional material was examined from Malaysia, Borneo, Myanmar, Java, Moluccas Islands, Philippines, West Papua, Papua New Guinea and several of the archipelagos east of Papua New Guinea. Several taxa may be represented across this broad range; however, I have chosen to refrain from describing additional species because of the extreme morphological variation and difficulty in distinguishing between various populations. Separate diagnoses could not be formulated for any of the populations studied. The genus is restricted almost exclusively to bat host species of Rhinolophus and Hipposideros and all specimens (when noted by collectors and/or removed in situ from alcohol preserved material) were removed from the wings with the exception of two specimens that were removed from the urogenital area (penis) of R. affinis . A preference for species of either Rhinolophus or Hipposideros could not be demonstrated, as species of each genus are commonly infested. It is unique (and possibly an incorrect record or a transcription error) that an ascodipterine was found on R. amplexicaudatus , a member of the family Pteropodidae (Old World fruit bats).
The origin of the subfamily Ascodipterinae appears to be the Afrotropical Region with subsequent easterly radiation to Asia and Australia. This is supported by the greater species diversity of Ascodipteron seen in the Afrotropical Region. Maabella evolved much later from Ascodipteron stock and is of recent origin. This evolutionary trend can be seen in the variable external morphology of this monotypic genus across its range.
The head (labial theca, gena, and lateral vertices) is similar across the range of this species; however, the size, abundance, and distribution of the setae differ. In general, the abundance of spiniform setae on the labial theca were greater among specimens from mainland Asia (SE China, Myanmar, Vietnam, Thailand, and Malaysia) than those from Borneo, Java, the Moluccas, Philippine Islands, West Papua, Papua New Guinea ( PNG), and archipelagos east of PNG; however, the labial thecae of some material from Malaysia and Myanmar (some from the same host specimens) had fewer and smaller spiniform setae. The number and length of setae were variable on the gena, the mesopleuron and the pteropleuron. Material from Java, the Islands in the Moluccas, and Borneo tend to have shorter and less numerous setae on these structures than specimens from all other areas. The chaetotaxy of the genital aperture was within the acceptable range of variation for the species (MSS: 1–5; VSS: 4–9; and genital plate: 1–3, usually 3). Setae in the R5 were identical for specimens throughout the region, with the exception of specimens from Java, which were vestigial or stump-like. The apical setae on trochanter were truly parallel only in Chinese populations, whereas the setae were more divergent and generally fewer in number in other populations, especially those from the oceanic islands. These differences could not be attributed to different host species; however, the variability of so many characters in this monotypic genus would imply that speciation is actively occurring among different populations parasitizing several rhinolophid host genera and species.
| BPBM |
Bishop Museum |
| V |
Royal British Columbia Museum - Herbarium |
| ROM |
Royal Ontario Museum |
| RMNH |
National Museum of Natural History, Naturalis |
| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
| AMNH |
American Museum of Natural History |
| FMNH |
Field Museum of Natural History |
| BYU |
Monte L. Bean Life Science Museum |
| BM |
Bristol Museum |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |