
Talitroides alluaudi ( Chevreux, 1896 )
|
publication ID |
https://doi.org/10.1080/00222933.2016.1180719 |
|
DOI |
https://doi.org/10.5281/zenodo.4328629 |
|
persistent identifier |
https://treatment.plazi.org/id/C12FB813-FFAC-FFE8-FEDC-FEBEFBA4FF2C |
|
treatment provided by |
Carolina |
|
scientific name |
Talitroides alluaudi ( Chevreux, 1896 ) |
| status |
|
Talitroides alluaudi ( Chevreux, 1896) BOLD:ACH9326
As Talitrus alluaudi Chevreux 1896, p 112 ; Talitroides alluaudi Chevreux, 1896 , Palmén 1949, p 61; Morino and Ortal 1993, p 332.
Material examined
4A, Hungry Bay mangrove forest in leaf litter; 5A, Spanish Point on limestone cliff, under terrestrial shrub leaf litter; 11A, near the same location as 5A.
Distribution
Only coastal locations incidental to the island-wide, supralittoral survey were sampled, so we are unable to describe the general distribution of this taxon in terrestrial habitats of Bermuda.
Remarks
The smallest talitrid at < 7 mm body length. This terrestrial taxon is probably common throughout the island where damp, well-rotted humus is found under leaf litter.
Key for the talitrids of Bermuda
1. Immature and adults < 7 mm body length, second antenna <25% of body length ............................................................................................................................ Talitroides alluaudi Immature and adults> 7 mm body length, second antenna> 25% of body length 2
2. Male second gnathopod dactylus with blunt tip and incised propodus palm; second antenna peduncle much enlarged in older males................................................................. 3 Male second gnathopod dactylus with fine tip and smooth propodus palm; second antenna peduncle not, or only slightly, enlarged in male................................................. 4
3. Male adult peraeopod 7 merus and carpus not incrassate; ovigerous female in oostegite 5 with equal numbers of fine setae on anterior and posterior margins .................................................................................................................. Platorchestia monodi View in CoL Male adult peraeopod 7 merus and carpus incrassate; ovigerous female with more fine setae on the anterior than the posterior margin of oostegite 5................................. ............................................................................................................................. Platorchestia platensis View in CoL
4. Dorsal pigment patterns on anterior peraeon absent; distolateral robust seta present on first uropod peduncle; four to seven tufts of ‘ comb ’ setae on male propodus of peraeopod 7............................................... ............................................... Tethorchestia antillensis View in CoL Dorsal pigment patterns conspicuous ( Figure 2 View Figure 2 ); distolateral robust seta absent on first uropod peduncle; maximum of four tufts of ‘ comb ’ setae on male propodus of peraeopod 7.......................................................................... Mexorchestia carpenteri carpenteri View in CoL
N.B. The few distinguishing features between the two Platorchestia View in CoL species are based on LeCroy (2011), and this reference should be consulted. Only adult males and females can be identified morphologically.
The formalin-preserved specimens, now in 50% isopropanol, are archived at the Bermuda Aquarium, Museum and Zoo (BAMZ) at Flatt ’ s Village, Bermuda, with the exception of two samples (8C archived at the Atlantic Reference Centre, Huntsman Marine Science Centre, St Andrews, N.B., Canada, and 16B formalin sample lost during sorting). Ethanol-preserved specimens, with the exception of those few consumptively analysed for DNA extraction, are archived at the Biodiversity Institute of Ontario, University of Guelph, Ont., Canada
Natural history
One feature of the Bermuda coastline is the relatively few beaches available where wrack and driftwood can strand. The popular South-shore beaches are cleaned of wrack in the summer months for the benefit of tourists. In winter months wrack removal is less frequent and, as pointed out by Healy and Coates (2003), this provides an opportunity for animals, including talitrids, to become established. Most of the coastline consists of a vertical, limestone, eroding cliff- face from a few to 30 m in height above sea level, where seaborne drifting materials are unable to strand. Nineteen beach locations were investigated, most < 50 m in linear length, which were spread throughout Bermuda ( Figure 2 View Figure 2 ). The name and co-ordinates of each sampling location are further specified in Table 1.
The following marine and terrestrial ecotopes harbouring talitrids were recognized in Bermuda:
1. Supralittoral, semi-tropical wrack on sand ( Table 1). The wrack consisted principally of Sargassum sp., sometimes mixed with other macroalgae, such as Enteromorpha sp., and sometimes with leaves of native terrestrial shrubs, such as the buttonwood, Conocarpus erectus L., and the bay grape, Coccolobo sp. At location 5B ( Figure 2 View Figure 2 and Table 1), which contained both P. monodi and M. c. carpenteri , the wrack was principally of senescent, bleached Enteromorpha sp., with Sargassum sp., also present. The wrack at Coot Pond (14B in a small inland sea), with both P. monodi and Tethorchestia antillensis , consisted mostly of leaves of land plants with a little Sargassum sp. The upper littoral of the inland sea at Coot Pond was colonized by unidentified woody shrubs. From a total of 19 supralittoral locations sampled, 13 were mostly of limestone sand. At Lagoon Park (16B) a small bivalve midden (valves of Arca zebra ) with buried Sargassum and terrestrial plant leaves, a few P. platensis BOLD:AAA2949 and P. monodi BOLD:AAB3402 were found.
2. Supralittoral, semi-tropical wrack on a rocky shore ( Table 1). The shore at Mullet Bay (8A) consisted of small limestone rocks with very little wrack. At Blue Hole (12B), the station sampled was close to the road causeway and was of mixed sand and small limestone rocks with small amounts of Sargassum wrack, making sampling difficult. At Devil ’ s Hole (12C), in the inland lagoon of Harrington Sound, the rocky shore was abbreviated by a wall with the wrack being of rubbish and terrestrial plant leaves. At Tobacco Bay Park (14A) on the seaward side of the coastline, copious Sargassum wrack mixed with anthropogenic litter was present on a rocky shore. At the Lagoon (15B), a small inland bay with seawater exchange in a pipe passing under the road, a mixed sand/rocky shore had only terrestrial plant wrack. At Lagoon Park (16A) the upper shore consisted of large limestone rocks on which wrack and driftwood were drying, with no talitrids found. A total of six of the locations sampled had a rocky or mixed sand/rock substratum, and P. monodi BOLD:AAB3402 was found in Devil ’ s Hole (12C) and a mixed sample at Tobacco Bay Park (14A), as shown in Table 1.
3. Supralittoral, semi-tropical driftwood ( Table 1). No large tree trunks or whole trees were found during the fieldwork. The largest piece (~ 5 m length) was a branch at 4B with no evidence of talitrids, or other inhabitants, with hard consistency and no evidence of rotting. On the sandy shore at 4B mostly occupied by Sargassum wrack, two coconut husks were opened, with no talitrids present. A small log (~ 1 m long) was found at 7A without talitrids. At Bailey ’ s Bay (8B) a small piece of driftwood (~ 30 × 15 × 5 cm) had obviously been at sea for an extended period as evidenced by the large holes ( Figure 4 View Figure 4 ) cut by the marine boring bivalve Teredo sp. A small wood sample from this was identified by Dr Peter Gasson of the Royal Botanic Gardens at Kew, UK, as spruce ( Picea sp.) and hence not from Bermuda. Two slow-moving talitrids were retrieved from this sample and proved to be P. monodi BOLD:AAB3402. It is of interest that the talitrids collected from wrack on the sand beach at Bailey ’ s Bay (8C) were P. platensis BOLD:AAA2949. Small log samples were found at 12A ( 45 cm long), 14A ( 30 cm), and 16A (both 50 cm long), all of which lacked talitrids. At 16A and 16B a coconut husk at each was opened without finding talitrids. Similarly, a coconut with decomposing milk still present at 16B also did not contain talitrids. Hence, we found driftwood samples at only seven of the 19 locations sampled, with talitrids in only one sample. The P. monodi BOLD: AAB3402 present were clearly in a secondary habitat ( Figure 4 View Figure 4 ), and no driftwood specialist talitrids were found.
4. Terrestrial, semi-tropical, coastal, forest leaf litter. The microhabitat in which T. alluaudi was found appeared to be superficially similar at each of the two locations (Hungry Bay mangrove forest in leaf litter and Stovell ’ s Bay on Spanish Point under shrubs on the limestone cliff) in which they were found, i.e. under terrestrial forest leaf litter where there was a dark-coloured, almost black humus layer. They were present with typical soil inhabitants, such as insect larvae, centipedes, millipedes, isopods, ants and also two individuals of P. monodi BOLD AAB3402.
Molecular data
From the 80 specimens selected for generating novel sequences for this study, only 63 talitrid specimens were successfully amplified. The failures included nine specimens of Platorchestia sp. and all eight specimens attempted for Tethorchestia antillensis ; therefore, we were only able to recover molecular data from four of the five morphologically defined species that were found in Bermuda. The strong taxon-specific pattern of polymerase chain reaction failure for Tethorchestia antillensis suggests substitution in COI in one of the primer binding regions, which requires further study. Most of the COI sequences included the full length of the barcode region (658 base pairs), but a few were slightly shorter.
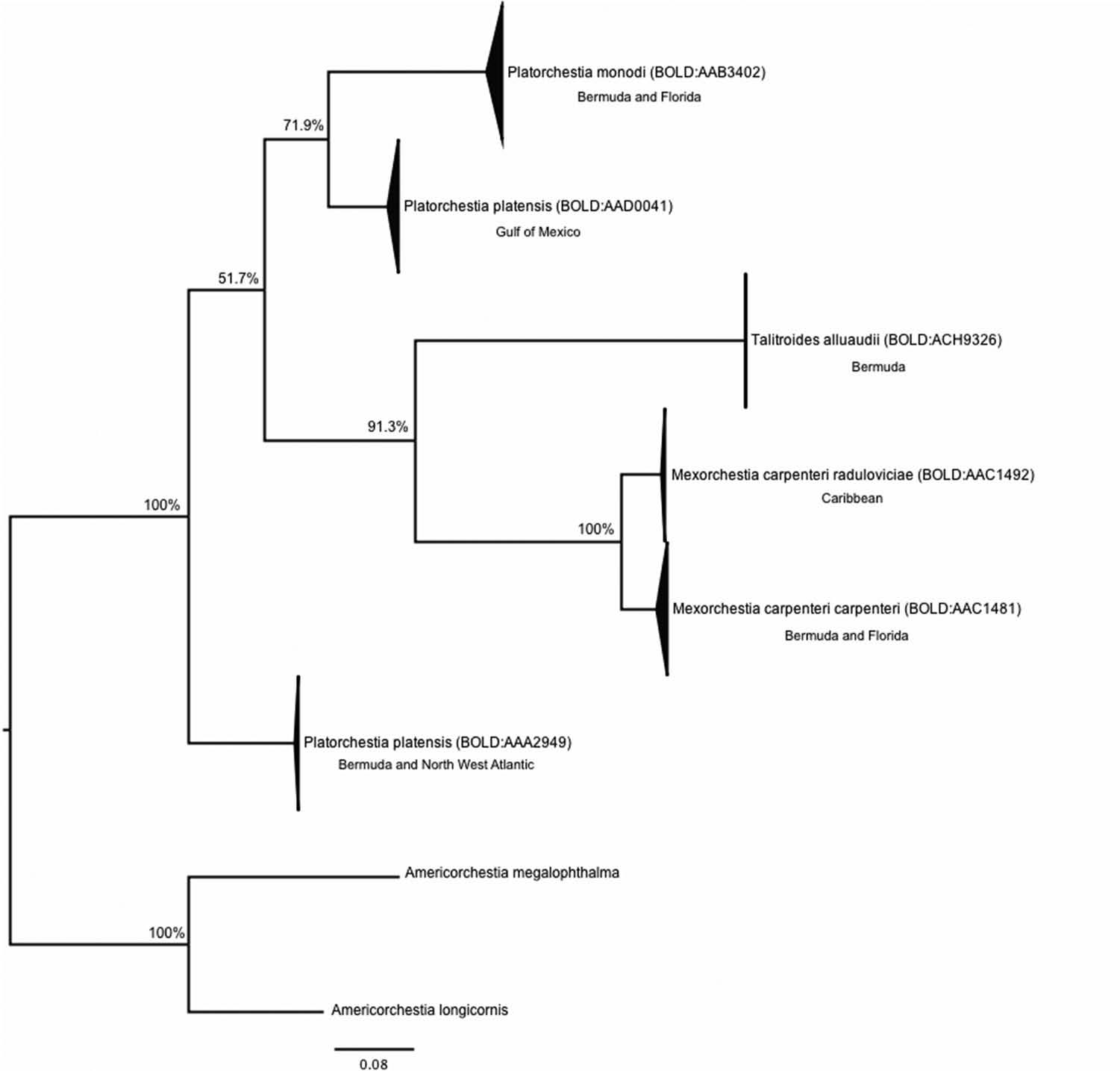
DNA barcoding of talitrids of Bermuda showed four BINs, corresponding to four morphological species. Specimens from M. c. carpenteri (BOLD:AAC1491), T. alluaudi (BOLD:ACH9326), P. monodi (BOLD:AAB3402) and P. platensis (BOLD:AAA2949) were shown to be each monophyletic with other members of their species in a maximum likelihood tree ( Figure 5 View Figure 5 ). The clade of P. monodi was formed by individuals from Bermuda and Florida, whereas the clade of P. platensis contained specimens from both Bermuda and the North Atlantic coast.
Haplotype diversity
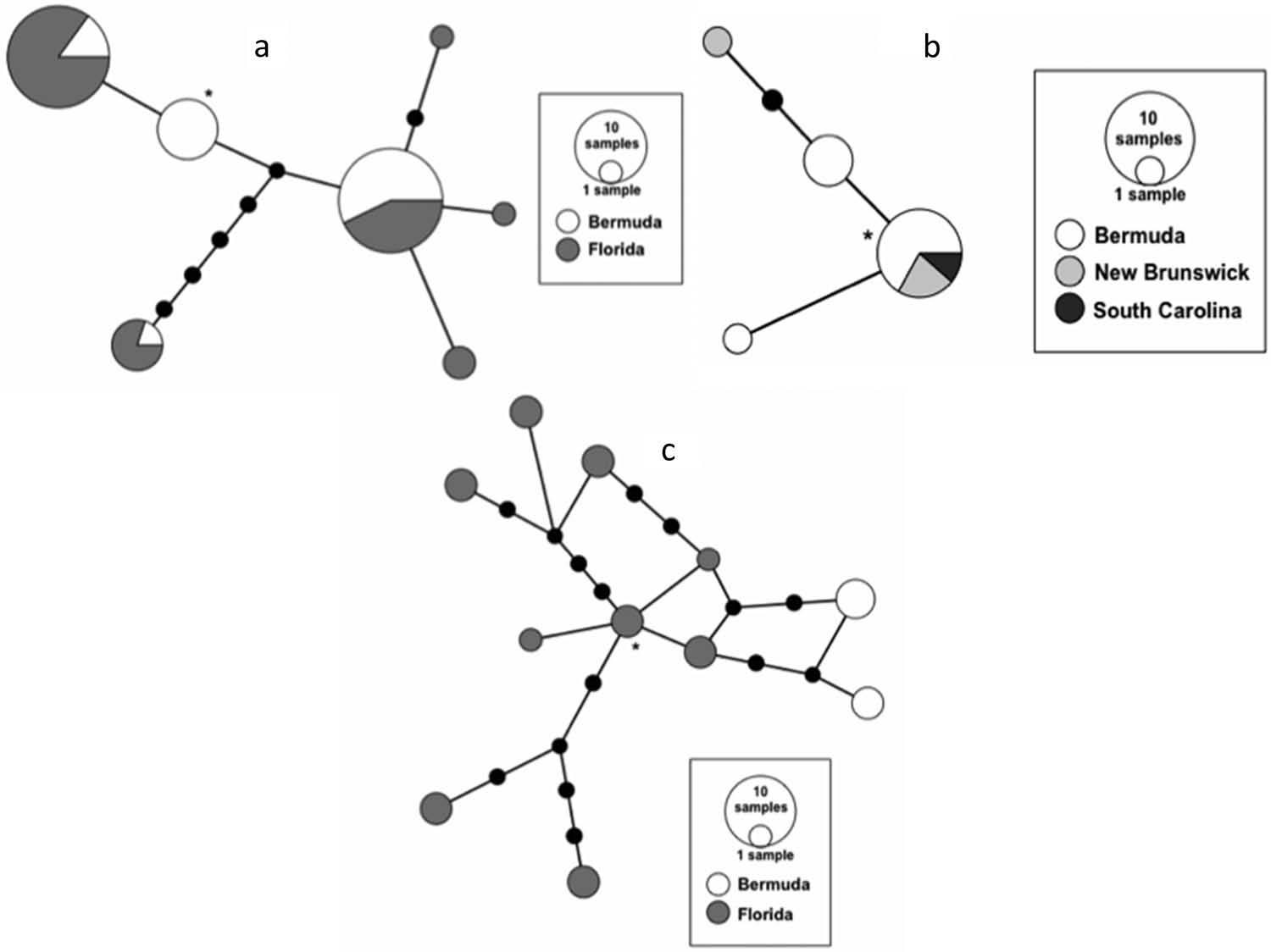
Haplotype networks were created to explore the haplotype distribution for three BINs found in Bermuda and other coastline localities from North America/Gulf of Mexico.
Seven haplotypes were identified for P. monodi BIN BOLD:AAB3402 among the 57 individuals sequenced. Three haplotypes are shared between the Florida and Bermuda localities. However, the other four haplotypes are only present in one of the two populations. One of the shared haplotypes is six or more mutational steps from all other haplotypes. Haplotype distribution by locality is presented in the median-joining network ( Figure 6 View Figure 6 ), where the ancestral haplotype according to the TCS analysis is marked with an asterisk. The median-joining network suggested that the presumptive ancestral haplotype is present only in Bermuda ( Figure 6 View Figure 6 ).
For P. platensis BIN BOLD: AA2949 , four haplotypes were identified among the 14 specimens assigned to this BIN ( Figure 6 View Figure 6 ). Only one haplotype, which has also been identified as the presumptive ancestral haplotype, is shared between Bermuda, South Carolina and New Brunswick localities . The other three haplotypes are unique for Bermuda or New Brunswick localities .
On the other hand, 11 haplotypes were identified for M. c. carpenteri ( BOLD: AAC1491 ) from among the 21 specimens sequenced from this BIN ( Figure 6 View Figure 6 ) . However, none of these haplotypes are shared between Florida and Bermuda localities. Two haplotypes are present in Bermuda, and are three or more mutational steps away from all Florida haplotypes, whereas the rest are only present in Florida. For this clade the presumptive ancestral haplotype occurs only in Florida, which is home to a population that is much more genetically diverse than that in Bermuda ( Figure 6 View Figure 6 ).
Population genetics analyses were not performed for BINs BOLD:AA2949 and BOLD: AAC1491, due to small sample size, but were performed for P. monodi BIN BOLD: AAB3402. The AMOVA suggests a significant degree of heterogeneity among populations with an FST = 0.336 ( p <0.001) ( Table 2). Pairwise comparison between the two populations of P. monodi , when also considering haplotype distances, are consistent with the AMOVA results (corrected average pairwise difference = 0.3374, p = 0.00000). The results from AMOVA and pairwise comparison between populations revealed that most genetic variation was observed within (66.4%) rather than between (33.6%) popu- lations. The Tajima ’ s D -tests for Bermuda (Tajima ’ s D = −0.87506) and Florida (Tajima ’ s D = 0.10341) populations were not significant ( Bermuda p = 0.195, Florida p = 0.605), which is consistent with a neutral model of COI molecular evolution in this data set.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Talitroides alluaudi ( Chevreux, 1896 )
| Wildish, D. J., Smith, S. R., Loeza-Quintana, T., Radulovici, A. E. & Adamowicz, S. J. 2016 |
Talitrus alluaudi
| Chevreux 1896: 112 |
Talitroides alluaudi
| Chevreux 1896 |