
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/ 10.7717/peerj.6573 |
|
DOI |
https://doi.org/10.5281/zenodo.3808813 |
|
persistent identifier |
https://treatment.plazi.org/id/C34887A8-FF99-FFEE-FDB4-AD048E6962B7 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
Here we report on the presence of feeding traces on the caudal vertebra of a hadrosaurid dinosaur (BMR P2007.4.1, ‘‘Constantine’’). Based on the shape and orientation of the traces, and the known fauna of the Hell Creek Formation, they are interpreted to be feeding traces of a large theropod dinosaur, such as Tyrannosaurus rex ( Erickson & Olson, 1996; Horner, Goodwin & Myhrvold, 2011). By comparing the dimensions and spacing of the traces with the maxillae and dentaries of specimens of Tyrannosaurus rex of different ontogenetic stages, we interpret these tooth marks to be feeding traces from a juvenile Tyrannosaurus rex and discuss the insights the specimen provides for juvenile tyrannosaur feeding behavior.
Geologic setting
Specimen BMR P2007.4.1 is a partial hadrosaurid skeleton collected from the Upper Cretaceous Hell Creek Formation of Carter County, southeastern Montana in the Powder River Basin ( Fig. 1 View Figure 1 ). This specimen was collected on public lands under BLM Permit # M96842 View Materials -2007 issued to Northern Illinois University and is accessioned at the Burpee Museum of Natural History in Rockford, IL. Exact coordinates for the location are on file in the paleontology collections at the Burpee Museum of Natural History (BMR), where the specimen is reposited.
The collection locality is composed of a 4 m fine-grained, gray-tan lenticular sandstone within a larger surrounding blocky mudstone unit ( Fig. 2 View Figure 2 ). The sandstone lacks bedforms, resulting from either (a) rapid accumulation (resulting in a lack of sedimentary structures), or (b) sedimentary structures that were obliterated by later currents or bioturbation, and is rich in rounded and weathered microvertebrate remains. The site is stratigraphically positioned approximately 44 m above the underlying Fox Hills–Hell Creek contact
and overlies 0.5 m of siderite, which sits above a 5 m blocky mudstone. Grains are subrounded to subangular. Microvertebrate and fragmented macrovertebrate fossils are abundant and heavily rounded and abraded ( Peterson, Scherer & Huffman, 2011). The fine-grained composition suggests a channel-fill deposit, overlying a floodplain deposit ( Murphy, Hoganson & Johnson, 2002; Peterson, Scherer & Huffman, 2011). The taphonomic distribution of the elements and their stratigraphic position suggests the skeleton was subaerially exposed on a floodplain for a considerable period of time prior to burial, allowing for weathering, disarticulation, and removal of many skeletal elements.
MATERIALS & METHODS
Specimen BMR P2007.4.1 consists of weathered pelvic elements (sacrum, left and right ilia), three dorsal vertebrae, and two proximal caudal vertebrae ( Fig. 3 View Figure 3 , Table 1 View Table 1 ). The dorsal vertebrae were too weathered for collection, though their dimensions and relative locations within the quarry assemblage were measured and documented. Additionally, a series of heavily-weathered bone fragments and a small shed theropod tooth ( Saurornithoides sp.) were also collected.
The ilium of BMR P2007.4.1 possesses a number of hadrosaurid characters such as (1) a shallow morphology, (2) a ∼ 23 ◦ preacetabular process in medial view relative to the main body, and (3) a well-developed supra-acetabular process caudal to the acetabulum. While these characters are common among hadrosaurids, the stratigraphic position of BMR P2007.4.1 suggests it is attributable to the Late Cretaceous hadrosaurid Edmontosaurus (i.e., Brett-Surman & Wagner, 2007; Campione, 2014).
The centra of the two caudal vertebrae lack any evidence for hemal arch attachments, suggesting they are among the more cranial-positioned caudal vertebrae, such as C1–C4 ( Campione, 2014). One of the caudal vertebrae possesses three v-shaped indentations on the ventral surface of the centrum ( Figs. 4A–4E View Figure 4 ; Figs. S1 and S2 View ). These traces feature collapsed cortical bone within the indentation, producing puncture marks (sensu Binford, 1981). The punctures penetrate 5 mm deep, are spaced 68 mm apart from their apical centers, show no signs of healing, and are inferred to have been created postmortem as feeding traces (e.g., Noto, Main & Drumheller, 2012; Hone & Tanke, 2015; McLain et al., 2018). The v-shape preserved in each puncture indicates that the original teeth would have possessed a prominent keel, though no striations from serration marks are present in the traces (sensu D’Amore & Blumensehine, 2009).
The large size and shape of the punctures suggests that they were produced by a largeto medium-bodied carnivore. Such carnivores from the Hell Creek Formation include tyrannosaurs such as Tyrannosaurus rex ( Erickson & Olson, 1996; Horner, Goodwin & Myhrvold, 2011), medium-sized dromaeosaurids such as Dakotaraptor steini ( DePalma et al., 2015) , and crocodylians such as Borealosuchus sternbergii , Brachychampsa montana , and Thoracosaurus neocesariensis ( Matsumoto & Evans, 2010) . By comparing the shape and orientation of the traces to the teeth of these carnivores from the Hell Creek Formation, they are hypothesized to be bite marks from a large theropod dinosaur, such as Tyrannosaurus rex ( Erickson & Olson, 1996); crocodylian teeth are circular in cross-section and too small, and dromaeosaurid teeth—even large dromaeosaurids such as D. steini —are too small and laterally-compressed to have produced the punctures observed on BMRP2007.4.1.
To test this hypothesis, the punctures on the caudal vertebra of BMR P2007.4.1 were first coated in ReboundTM 25 platinum-cure silicone rubber (Smooth-On) in order to make a silicone peel of the punctures in order to better visualize the morphology and dimensions of the teeth responsible for the traces ( Figs. 5A–5B View Figure 5 ; Figs. S3 and S4 View ). These ‘‘teeth’’ were then compared with the dental dimensions and spacing of two Tyrannosaurus maxillae and dentaries. To approximate the ontogenetic stage of the tyrannosaur, a late-stage juvenile specimen ( BMR P2002.4.1, ‘‘Jane’’ ) histologically determined to be approximately 11–12 years old at the time of death ( Erickson et al., 2006) that possesses laterally compressed, sharp crowns, and a mature specimen ( BHI 3033, ‘‘Stan’’ ) with robust, blunt crowns were utilized.
All specimens were digitized via triangulated laser texture scanning with a NextEngine 3D Laser Scanner, capturing data at seven scanning divisions in high-definition (2.0k points/in 2). The resulting digital models were built with the NextEngine ScanStudio HD Pro version 2.02, and finalized as STL models ( Figs. S1–S8 View ). Scanning was conducted at the Department of Geology at the University of Wisconsin-Oshkosh in Oshkosh, WI.
The tooth spacing of both adult and late-stage juvenile tyrannosaur maxillae and dentaries were measured for both immediately-adjacent teeth and teeth from alternating replacement positions (i.e., Zahnreihen), and compared with the spacing of the punctures ( Figs. 6A–6B View Figure 6 ), similar to Fahlke ’s (2012) investigation of likely Basilosaurus bite marks on specimens the smaller whale Dorudon . Furthermore, the cross-sectional morphology
of adult and late-stage juvenile tyrannosaur maxillae and dentaries were measured labiolingually and mesiodistally at a 5 mm apical depth for each tooth crown, and plotted with measurements from the punctures found on BMR P2007.4.1 ( Fig. 7 View Figure 7 ).
RESULTS
The mesiodistal width measurements from the silicone peel taken from BMR P2007.4.1 average 7.8 mm and the labiolingual depth average was 5.2 mm. Maxillary and dentary teeth of the adult Tyrannosaurus ( BHI 3033 ) were found to be too large and widely spaced to have produced the punctures ( Figs. 7 View Figure 7 , 8A and 8B View Figure 8 ; Figs. S4 and S5 View ; Tables 2A–2C View Table 2 ). For BHI 3033 , the average dentary tooth crown mesiodistal width at 5 mm depth was 7.13 mm, and the average dentary tooth crown labiolingual depth at 5 mm was 4.10 mm. The average maxillary crown mesiodistal width at 5 mm were 7.72 mm, and the average maxillary crown labiolingual depth at 5 mm averaged to 4.21 mm.
However, the teeth of BMR P2002.4.1 produced similarly shaped punctures at 5 mm apical depth ( Figs. 7 View Figure 7 and 9 View Figure 9 ; Figs. S7 and S8 View ; Tables 2B–2C View Table 2 ). The puncture measurements taken from the peel, BMR P2007.4.1 demonstrate a mesiodistal width and labiolingual depth consistent with the measurements taken from the maxillary and dentary teeth of the late-stage juvenile Tyrannosaurus . When plotted against the mesiodistal width and labiolingual depth of the maxillary teeth, measurements from the peel taken from BMR P2007.4.1 fall well within the cluster radius created by the late-stage juvenile Tyrannosaurus , BMR P2002.4.1 ( Fig. 7 View Figure 7 ). Furthermore, the inferred crown spacing of the punctures closely matched those of the late-stage juvenile tyrannosaur maxilla ( Tables 3A–3B View Table 3 ).
DISCUSSION AND CONCLUSIONS
While feeding traces and bite marks attributed to mature tyrannosaurids are welldocumented in common Late Cretaceous taxa such as hadrosaurids and ceratopsians (i.e., Fiorillo, 1991; Erickson et al., 1996; Erickson & Olson, 1996; Jacobsen, 1998; Farlow & Holtz, 2002; Fowler & Sullivan, 2006; Peterson et al., 2009; Bell & Currie, 2009; Fowler et al., 2012; DePalma et al., 2013; McLain et al., 2018), the identification of juvenile tyrannosaur feeding traces adds insight into the role of juvenile theropods in Cretaceous ecosystems. The dimensions and spacing of the punctures closely matches the maxillary teeth of BMR P2002.4.1 , a late-stage juvenile (11–12 yr old) tyrannosaur which incidentally itself possesses morphologically similar craniofacial lesions previously interpreted as a conspecific bite ( Peterson et al., 2009).
Longrich et al. (2010) reported on evidence of cannibalism in T. rex based on a number of bitten and scored remains of Tyrannosaurus rex , some attributed to juvenile or subadult individuals. However, many of these traces resemble the ‘puncture and pull’ bite marks that have previously been attributed to T. rex ( Erickson et al., 1996; Erickson & Olson, 1996), and also include furrows and scores (sensu Binford, 1981).
Correlating traces in bone, such as tooth marks, to specific taxa and ontogenetic stages usually requires direct comparisons (e.g., Peterson et al., 2009; Fahlke, 2012). However, in cases where direct comparisons are not available, estimates can be made for tooth size, morphology, and spacing based on ontogenetic trajectories. While bite marks and feeding traces attributable to younger juvenile and hatchling tyrannosaurs have not yet been identified, the punctures present on the caudal vertebra of BMR P2007.4.1 provide direct evidence that late-stage juvenile Tyrannosaurus rex such as BMR P2002.4.1 possessed—at least in part—a similar diet as adults.
While bite marks resulting from active predation cannot easily be distinguished from postmortem feeding traces, the ventral position of the punctures in the caudal centrum of BMR P2007.4.1 suggests that the feeding was taking place postmortem with the hadrosaur already on its side ( Chure, Fiorillo & Jacobsen, 1998). The afflicted vertebra is from the cranial-most part of the tail. Observations of the feeding behaviors of carnivoran mammals and birds indicate that in most cases, consumption of the axial skeleton occurs after limbs and viscera have been consumed (e.g., Hill, 1980; Haglund, 1997; Carson, Stefan & Powell, 2000; Behrensmeyer, Stayton & Chapman, 2003). Hadrosaur tails had substantial muscles such as m. ilio-ischocaudalis and m. caudofemoralis longus ( Persons & Currie, 2014) that might be a target of early stage consumption. However, the ventral bite traces on BMR P2007.4.1 suggest that the tyrannosaur was feeding after the haemal complexes and most of the superficial hypaxial muscles and m. caudofemoralis longus had been removed. As such the punctures on BMR P2007.4.1 suggest later-stage carcass consumption and postmortem feeding behaviors.
The identification of penetrating bite marks attributable to not only Tyrannosaurus rex , but an individual of 11–12 years of age can potentially allow for the determination of the ontogeny of bite force in Tyrannosaurus rex and for comparison with other theropods (e.g., Barrett & Rayfield, 2006; Gignac et al., 2010; Bates & Falkingham, 2012). Studies on the estimated bite forces of an adult Tyrannosaurus rex have yielded a wide range of results. Estimates based on muscle volume proposed bite forces between 8,526 and 34,522 N, coupled with tooth pressures of 718–2,974 MPa, and a unique tooth morphology and arrangement to promote fine fragmentation of bone during osteophagy ( Gignac & Erickson, 2017). However, estimates incorporating likely muscle fiber length produced results over 64,000 N for adult T. rex ( Bates & Falkingham, 2018). Juvenile T. rex such as BMR P2002.4.1 have much narrower and blade-like tooth morphologies and were unlikely to have been able to withstand similar bite forces at this ontogenetic stage. Bates & Falkingham (2012) estimated a maximum bite force for BMR P2002.4.1 at 2,400–3,850 N, and hypothesized that an increase in bite force during growth could indicate a change in feeding behavior and dietary partitioning while approaching adulthood.
Observations on extant crocodylians have documented a wide variety of dietary partitioning during ontogeny (e.g., Tucker et al., 1996; Platt et al., 2006; Platt et al., 2013).
In the American crocodile ( Crocodylus acutus ), hatchling and small juveniles have a dietary overlap of over 80%, commonly feeding upon insects and crustaceans ( Platt et al., 2013). Alternatively, larger juveniles, subadults, and adults possess a dietary overlap of over 75%, consisting of more birds, mammals, fish, and other reptiles ( Platt et al., 2013). Comparable ontogenetic dietary partitions were also observed in Morelet’s crocodile ( Crocodylus moreletii ) ( Platt et al., 2006), and in Australian freshwater crocodiles ( Crocodylus johnstoni ) ( Tucker et al., 1996). However, crocodylians are less discriminant of food sources when scavenging (e.g., Antunes, 2017). While the punctures present on BMR P2007.4.1 are likely from postmortem scavenging behaviors of a juvenile tyrannosaur, the degree of dietary overlap or partitioning between juvenile and adult tyrannosaurs remains unresolved.
Despite not yet possessing the same feeding mechanisms of an adult Tyrannosaurus rex (i.e., bone-crushing and osteophagy), the punctures present on BMR P2007.4.1 demonstrate that late-stage juvenile and subadult tyrannosaurs were already biomechanically capable of puncturing bone during feeding, and were doing so without the large, blunt dental crowns of adults. Further identification of tyrannosaur feeding traces from different ontogenetic stages coupled with experimental studies of the biomechanics of tyrannosaur bite forces may reveal more insight into dynamic dietary trends and ecological role of Tyrannosaurus rex throughout ontogeny.





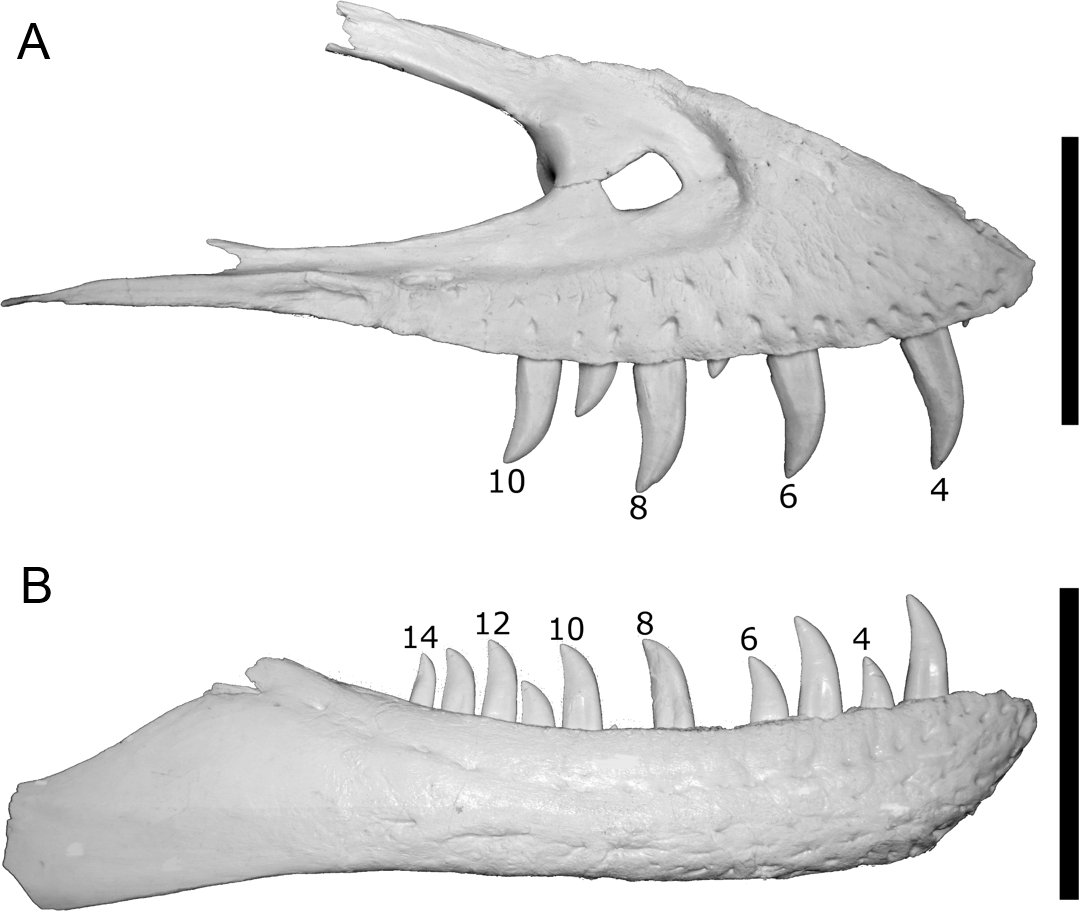
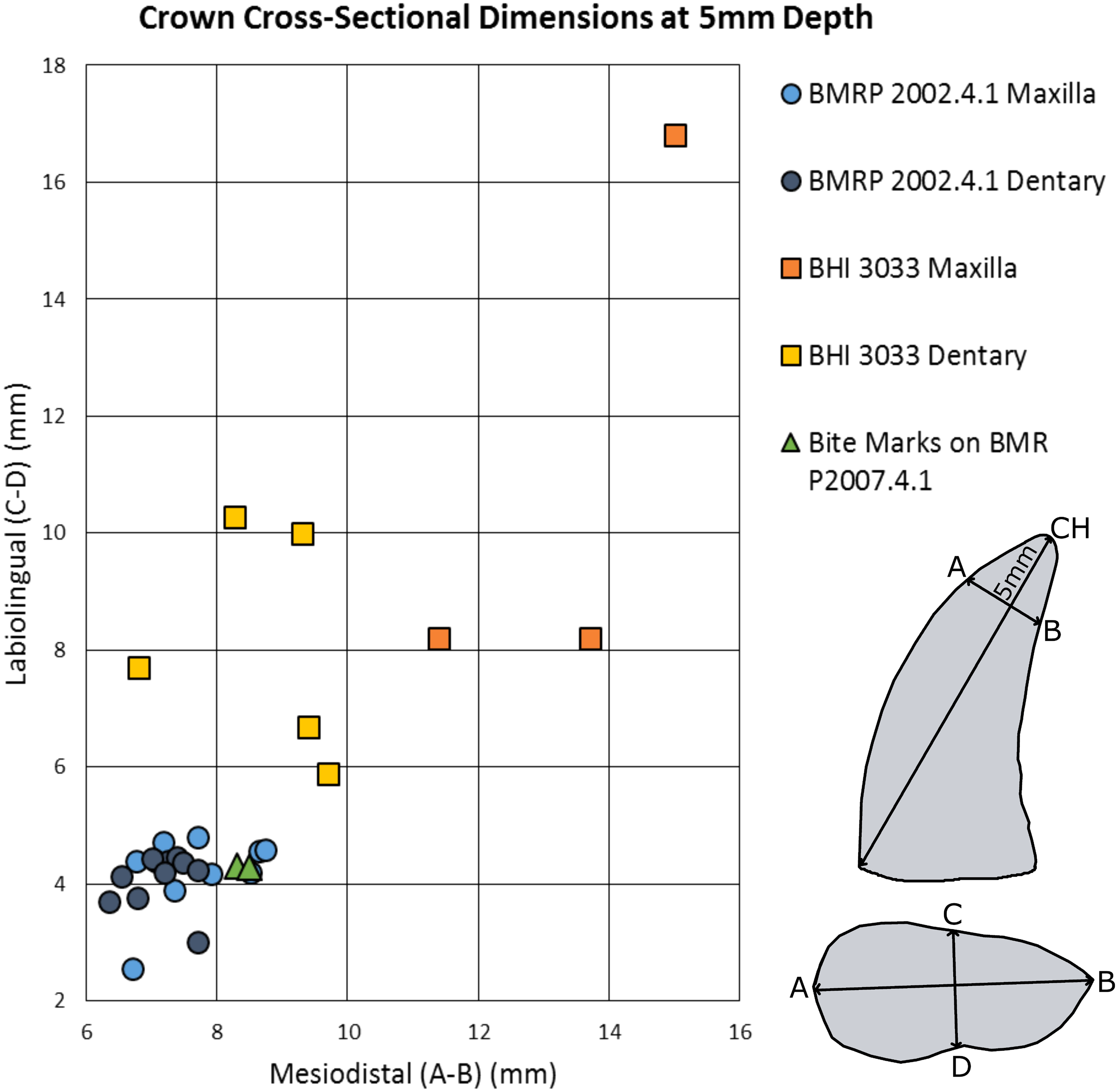
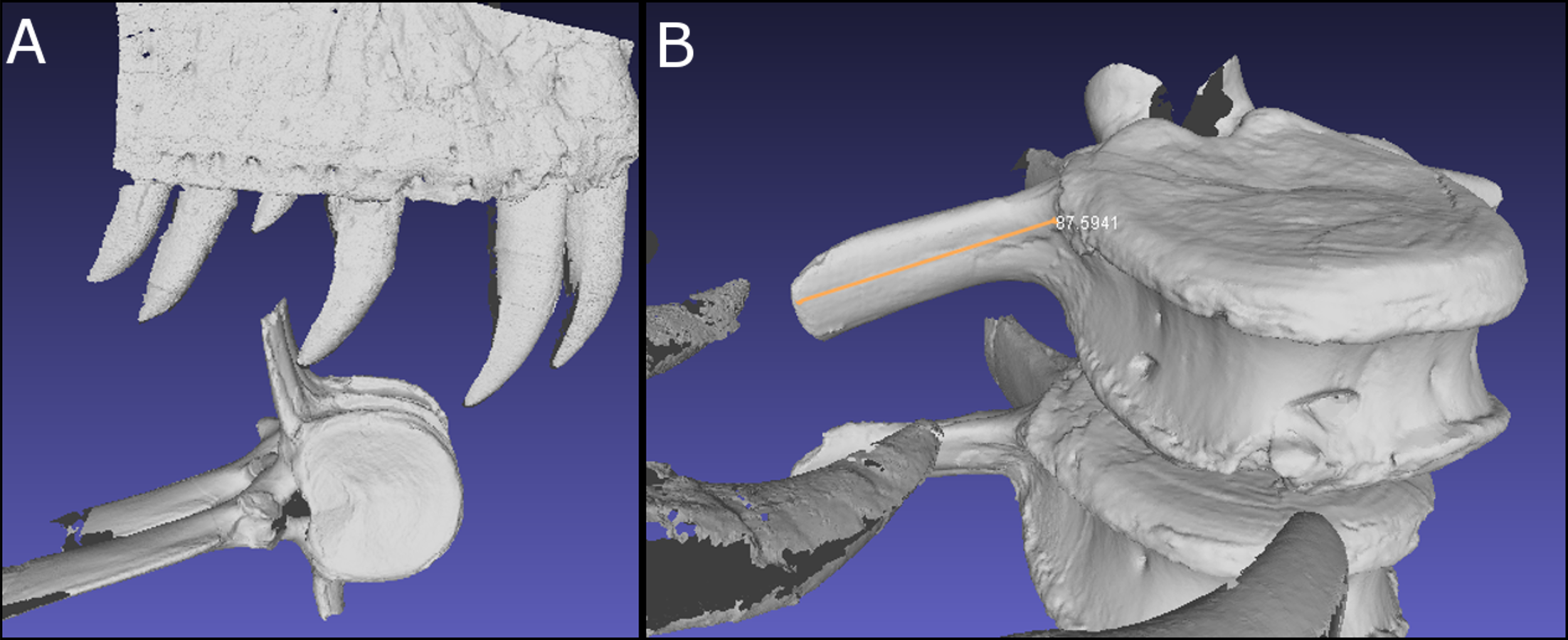

Table 1 Skeletal elements from BMR P2007.4.1. Recovered and recorded skeletal elements from the ‘‘Constantine Quarry’’ (BMR P2007.4.1) and taphonomic condition.
| Field number | Element | State/Condition |
|---|---|---|
| CON-2007-001 | Rib fragment | Abraded |
| CON-2007-002 | Left ilium | Heavily weathered |
| CON-2007-003 | Rib fragment | Abraded |
| CON-2007-004 | Sacrum and right ilium | Heavy to moderate weathering |
| CON-2007-005 | Neural arch | Fractured, but mild weathering |
| CON-2007-006 | Caudal vertebra | Mild weathering |
| CON-2007-007 | Caudal vertebra | Mild weathering |
| CON-2007-008 | Bone fragment | Heavily abraded |
| CON-2007-009 | Shed Saurornithoides sp. tooth | No apparent abrasion |
| CON-2007-010 | Dorsal vertebra | Heavily weathered, not collected |
| CON-2007-011 | Dorsal vertebra | Heavily weathered, not collected |
| CON-2007-012 | Dorsal vertebra | Heavily weathered, not collected |
Table 2 Measurements of tooth crowns of tyrannosaur specimens. Mesiodistal and labiolingual measurements of teeth at 5 mm depth from the crown apex for (A) BHI 3033, (B) BMR P2002.4.1, and (C) the inferred bite marks on BMR P2007.4.1. All measurements are in mm.
| A BHI 3033 | Maxilla | Dentary | ||
|---|---|---|---|---|
| Mesiodistal | Labiolingual | Mesiodistal | Labiolingual | |
| 15 | 16.8 | 9.3 | 10.0 | |
| 11.4 | 8.2 | 8.27 | 10.27 | |
| 13.7 | 8.2 | 6.8 | 7.7 | |
| 9.4 | 6.7 | |||
| 9.7 | 5.9 |
Table 3 Measurements of crown spacing in tyrannosaur specimens. Tooth crown spacing between maxillary (A) and dentary (B) teeth in the juvenile tyrannosaur BMR P2002.4.1. All measurements are in mm.
| A Crown spacing | Maxillary (mm) |
|---|---|
| 4–6 | 70.2 |
| 6–8 | 73.3 |
| 8–10 | 62.8 |
| Average | 68.7 |
| B Crown spacing | Dentary(mm) |
| 4–6 | 53.3 |
| 6–8 | 49.8 |
| 8–10 | 39.2 |
| 10–12 | 33.7 |
| 12–14 | 33 |
| Average | 41.8 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |