
Paraleucilla solangeae, Cavalcanti, Fernanda F., Menegola, Carla & Lanna, Emilio, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3764.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:66502B5F-5DBF-4C0C-A09A-6E4D6196D032 |
|
DOI |
https://doi.org/10.5281/zenodo.6134583 |
|
persistent identifier |
https://treatment.plazi.org/id/C4078793-FFD9-B73E-0ABA-A24A389CFB26 |
|
treatment provided by |
Plazi |
|
scientific name |
Paraleucilla solangeae |
| status |
sp. nov. |
Paraleucilla solangeae View in CoL sp. nov.
Etymology. For Dr. Solange Peixinho, in recognition of her dedication to the taxonomy of calcareous sponges, mainly species from Bahia and other states in northeastern Brazil.
Diagnosis. Paraleucilla with a single apical osculum ornamented by a short fringe formed by trichoxea and small sagittal tetractines. Tufts of diactines are positioned perpendicular to the sponge surface, crossing two thirds of the choanosome. They are regularly distributed ( i.e., equally spaced) along the sponge skeleton. The cortical skeleton is composed of giant tetractines. Both the outer and inner regions have a single type of sagittal triactines and tetractines. The atrium is lined with a tangential layer of sagittal triactines and tetractines.
Type material. Holotype: UFBA POR 3525 [Guarajuba ( 12°44’12.6’’S 38°05’12.4’’W), Camaçari, Bahia, Brazil; depth: 23 meters; --/ II/2008], Paratypes: UFBA POR 4242 [Guarajuba ( 12°44’12.6’’S 38°05’12.4’’W), Camaçari, Bahia, Brazil; depth: 23 meters; --/ II/2008], and UFBA POR 4243 [Guarajuba ( 12°44’12.6’’S 38°05’12.4’’W), Camaçari, Bahia, Brazil; depth: 23 meters; --/ II/2008].
Type locality. Guarajuba, Camaçari, Bahia, Brazil.
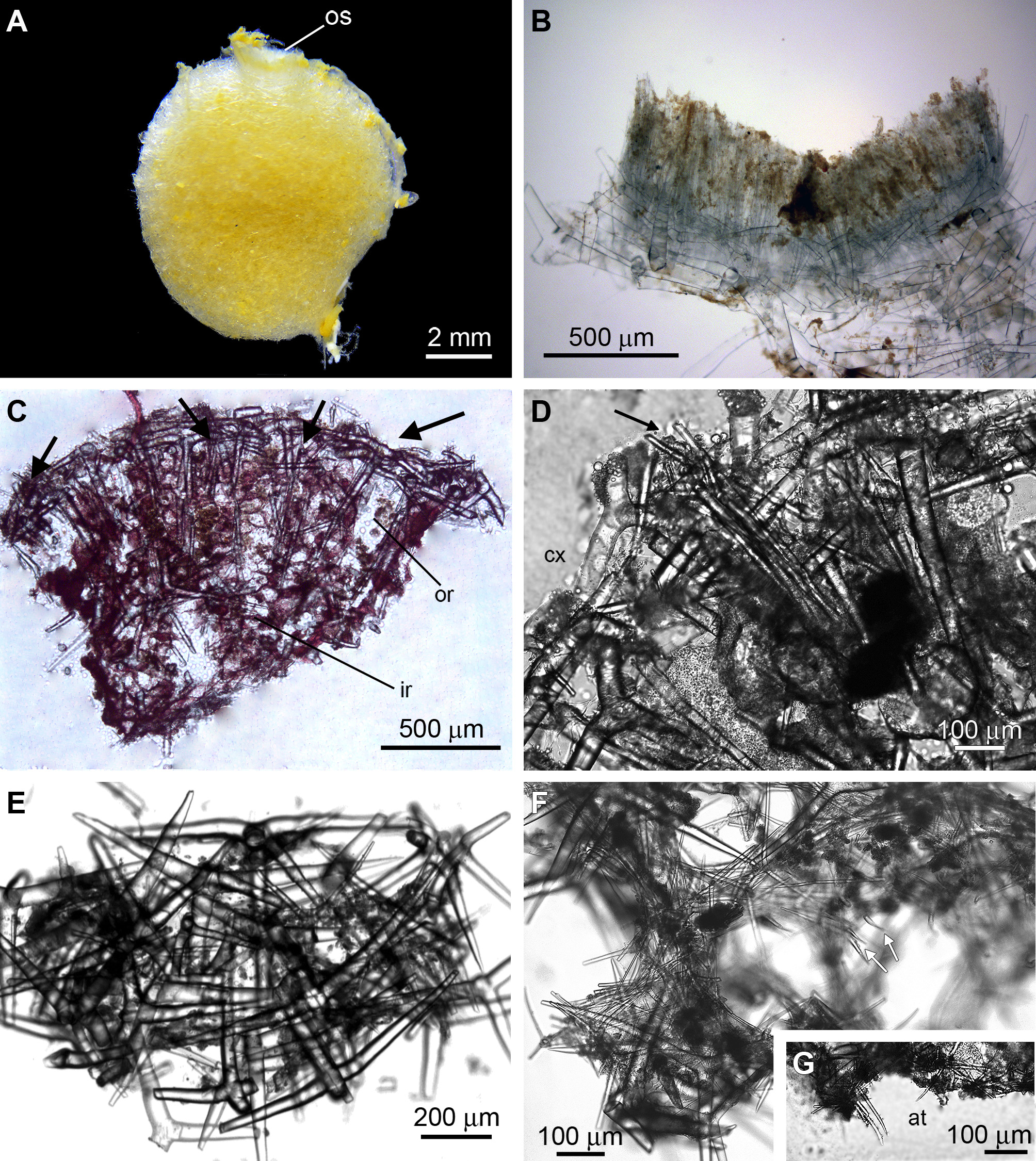
Description. The body shape varies from spherical to tubular. The holotype is the largest specimen, and it measures 8.0 x 8.0 mm (osculum-basis axis and width, respectively) ( Figure 2 View FIGURE 2 A). The color when alive is unknown, but the ethanol-preserved specimens are beige. The surface is smooth, but this is most likely because the tips of the diactines are broken (see description below). There is a single apical osculum, which in the holotype (UFBA POR 3525) and in one of the paratypes (UFBA POR 4242) is ornamented by a fringe of trichoxea ( Figure 2 View FIGURE 2 B). Small sagittal tetractines are present at the base of this fringe, and they are covered by the giant spicules that form the cortical skeleton. The body wall is thick in the holotype, which has the narrowest atrial cavity. The paratypes present a wide atrial cavity. The aquiferous system is leuconoid, with the rounded choanocyte chambers typical of the genus.
There are diactines perpendicular to the sponge surface, crossing two thirds of the choanosome (from the outer region to the inner region). They are organized in tufts that are regularly arranged in the sponge body ( Figures 2 View FIGURE 2 C, D). The cortical skeleton is composed of the basal system of giant tetractines ( Figures 2 View FIGURE 2 C, E), which point their apical actine toward the atrium. The inarticulated skeleton ( i.e., the outer region) is formed by these apical actines and by the unpaired actines of subatrial triactines and tetractines. Both categories are common in the subatrial skeleton, and they also form the inner region, where the spicules are irregularly distributed. The atrial skeleton is composed of tangential triactines and tetractines. The latter project their apical actines into the atrial cavity ( Figures 2 View FIGURE 2 F, G).
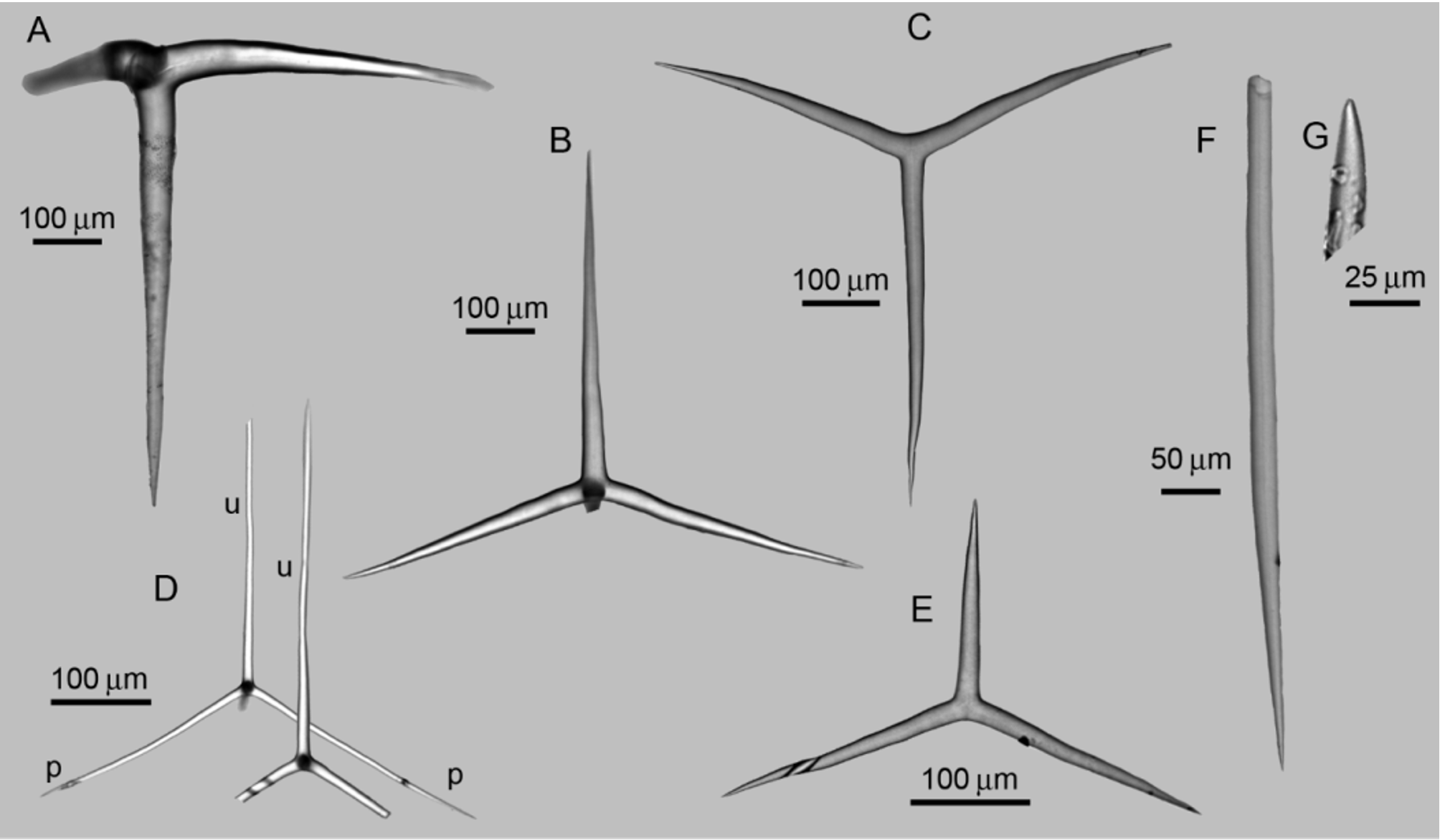
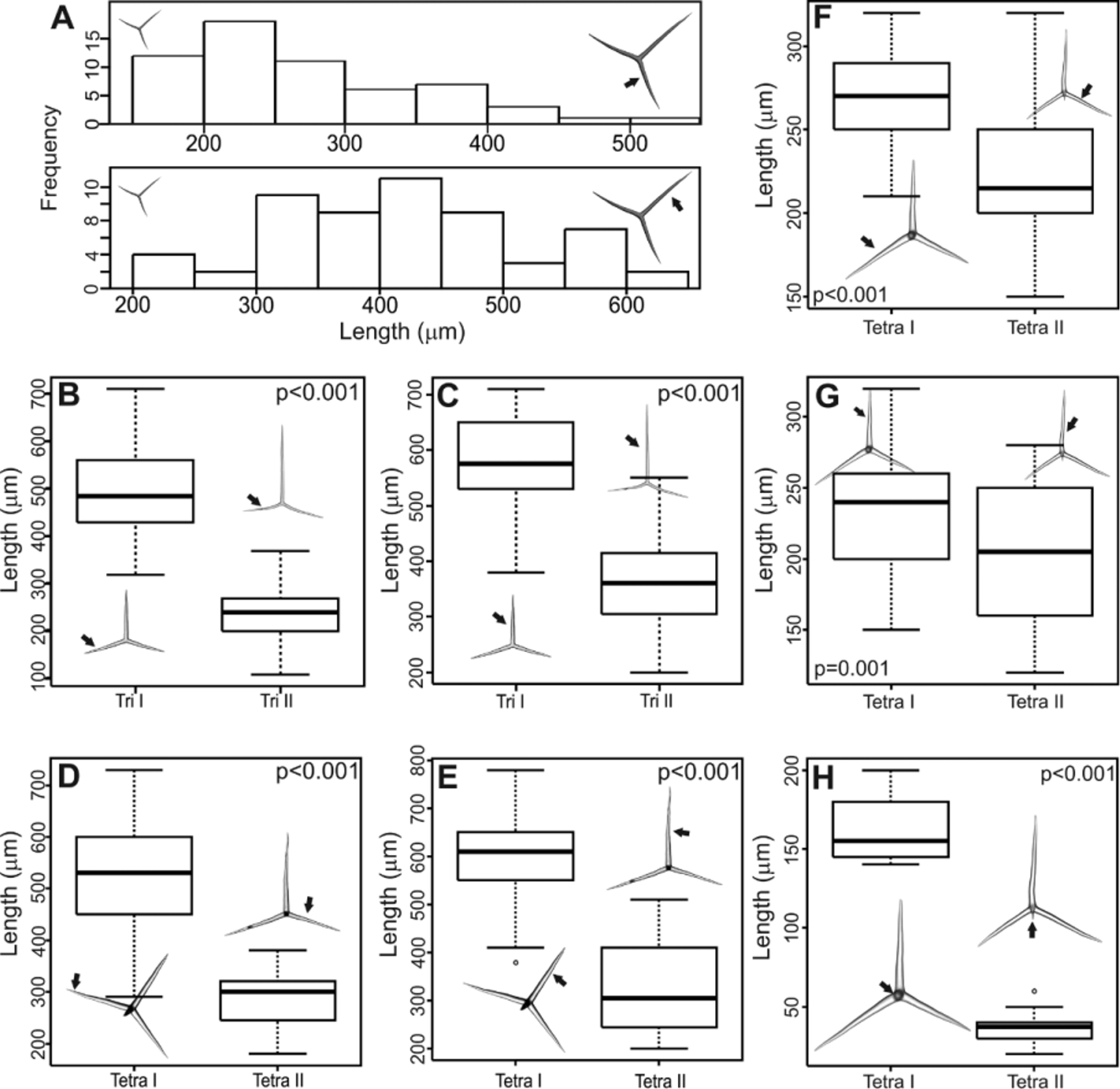
Spicules. ( Table 1). Cortical tetractines ( Figure 3 View FIGURE 3 A): Giant spicules. All actines are conical and have sharp tips. In the basal system, the unpaired actine is shorter than the paired actines, which are curved. The apical actine is straight. [Paired actines: 200 – 373.6 ± 27.8 – 570/ 35.4 ± 2.6 µm; unpaired actine: 190 – 311.2 ± 61.9 – 520.0/ 30.6 ± 6.2 µm; apical actine: 220 – 500.4 ± 48.2 – 750/ 36.8 ± 1.9 µm (n= 3 specimens)]. Subatrial tetractines ( Figure 3 View FIGURE 3 B): Sagittal. Actines are conical and have sharp tips. The paired actines are straight, whereas the unpaired actine is slightly undulated. In general, the latter is longer than the paired actines. The apical actine is short, smooth, and curved. [Paired actines: 160 – 359.6 ± 8.1 – 560/ 35.3 ±4.0 µm; unpaired actine: 170 – 403.3 ± 17.2 – 650/ 36.8 ± 4.2 µm; apical actine: 70 – 182.3 ±98.4 – 430.0/ 26.8 ± 6.1 µm (n= 3 specimens)]. Subatrial triactines ( Figure 3 View FIGURE 3 C): Sagittal. Actines are conical and have sharp tips. In general, the unpaired actine is longer than the paired actines, which are curved. In one of the paratypes (UFBA POR 4242), these spicules have a wide range of sizes. We confirmed that they comprise a single category by plotting a histogram ( Figure 4 View FIGURE 4 A). [Paired actines: 110 – 268.4 ± 57.0 – 550/ 16.5 ± 5.9 µm; unpaired actine: 170 – 412.4 ± 59.2 – 630/ 18.9 ± 5.8 µm (n= 3 specimens)]. Atrial tetractines ( Figure 3 View FIGURE 3 D): Thinner than any other spicule category. Actines are cylindrical and straight (with the exception of the apical one), and they have sharp tips. The apical actine is short and curved close to its tip. [Paired actines: 110 – 223.0 ± 30.7 – 350/ 9.0 ± 2.1 µm; unpaired actine: 100 – 260.0 ± 24.0 – 510/ 10.4 ± 0.4 µm; apical actine: 50 – 104.1 ± 14.6 – 170/ 9.1 ± 1.2 µm (n= 3 specimens)]. Atrial triactines ( Figure 3 View FIGURE 3 E): Sagittal, actines are conical and have sharp tips. The paired actines are curved and are longer than the unpaired one, which is straight. [Paired actines: 90 – 148.8 ± 7.4 – 280/10.5 ± 1.2 µm; unpaired actine: 40 – 88.6 ± 5.2 – 170/ 12.9 ± 0.6 µm (n= 3 specimens)]. Diactines ( Figures 3 View FIGURE 3 F, G): Straight and fusiform. Both tips are sharp, but whereas the proximal one (located inside the sponge) thins gradually ( Figure 3 View FIGURE 3 F), the distal tip (located outside the sponge) comes abruptly to a point ( Figure 3 View FIGURE 3 G). Frequently, this spicule has its distal tip broken. [160 – 498.0 ± 103.7 – 850/ 14.3 ± 2.8 µm (n= 3 specimens)].
Ecology. All the analyzed specimens were sampled by dredging during the monitoring of a submarine emission pipe on the North coast of Bahia (see Materials and Methods above). Specimen UFBA POR 3525 (the holotype) is attached to a fragment of an organism that is most likely a soft coral, and hydroids, calcareous algae, sand, and foraminiferans are also found on its basal surface. On the other specimens (the paratypes), only calcareous algae are observed.
Spicule/ Specimens Actine Length (µm) Width (µm) N
Mean SD Mean SD Cortical tetractines
UFBA POR 3525 (H) paired 381.3 75.3 38.3 9.9 30 UFBA POR 4242 (P) 396.7 82.3 33.5 8.9 30 UFBA POR 4243 (P) 342.7 96.0 34.3 9.4 30 UFBA POR 3525 (H) unpaired 267.5 36.0 26.2 4.8 0 4 UFBA POR 4242 (P) -- -- -- -- -- UFBA POR 4243 (P) 355.0 233.3 35.0 21.2 0 2 UFBA POR 3525 (H) apical 512.7 114.2 38.7 8.5 30 UFBA POR 4242 (P) 541.3 146.3 34.8 7.9 30 UFBA POR 4243 (P) 447.3 92.7 36.9 8.4 26 Subatrial tetractines
UFBA POR 3525 (H) paired 365.3 83.9 37.3 9.7 30 UFBA POR 4242 (P) 350.3 100.3 30.7 9.1 30 UFBA POR 4243 (P) 363.3 95.6 38.0 10.3 15 UFBA POR 3525 (H) unpaired 412.7 91.9 37.5 8.3 30 UFBA POR 4242 (P) 413.7 120.6 32.2 9.3 30 UFBA POR 4243 (P) 383.5 98.8 40.6 11.6 17 UFBA POR 3525 (H) apical 223.3 68.1 31.7 7.6 0 3 UFBA POR 4242 (P) 253.7 96.2 28.7 6.4 0 8 UFBA POR 4243 (P) 70.0 -- 20.0 -- 0 1 Subatrial triactines
UFBA POR 3525 (H) paired 248.7 67.1 15.5 4.8 30 UFBA POR 4242 (P) 332.7 77.1 22.8 6.8 30 UFBA POR 4243 (P) 224.0 84.4 11.2 3.6 20 UFBA POR 3525 (H) unpaired 382.7 97.2 17.7 6.0 30 UFBA POR 4242 (P) 480.7 81.7 25.2 6.6 30 UFBA POR 4243 (P) 374.0 77.1 13.7 3.7 30 Atrial tetractines
UFBA POR 3525 (H) paired 239.7 46.8 9.0 2.3 30 UFBA POR 4242 (P) 241.7 46.8 11.2 4.9 30 UFBA POR 4243 (P) 187.5 39.2 6.9 2.6 0 8 UFBA POR 3525 (H) unpaired 277.0 97.0 10.1 2.5 23 UFBA POR 4242 (P) 243.0 94.2 10.7 3.4 30 UFBA POR 4243 (P) -- -- -- -- -- UFBA POR 3525 (H) apical 87.9 24.4 7.8 2.6 29 UFBA POR 4242 (P) 116.3 21.6 10.3 3.2 30 UFBA POR 4243 (P) 108.2 34.0 9.1 7.3 11 Atrial triactine
UFBA POR 3525 (H) paired 144.0 30.2 11.0 2.6 20 UFBA POR 4242 (P) 157.3 36.0 11.3 3.7 30......continued on the next page Spicule/ Specimens Actine Length (µm) Width (µm) N
Mean SD Mean SD
UFBA POR 4243 (P) 145.0 30.4 9.1 7.3 11 UFBA POR 3525 (H) unpaired 90.0 32.4 13.0 3.4 20 UFBA POR 4242 (P) 93.0 34.8 13.5 3.5 30 UFBA POR 4243 (P) 82.8 23.0 12.3 7.8 30 Diactines*
UFBA POR 3525 (H) 499.6 129.3 15.2 3.6 30 UFBA POR 4242 (P) 601.0 111.5 16.5 3.9 30 UFBA POR 4243 (P) 393.5 87.8 11.1 4.5 14 Remarks. Paraleucilla solangeae sp. nov. is morphologically similar to other species of the genus Paraleucilla occurring in the SW Atlantic along the South American coast. It shares with P. perlucida Azevedo & Klautau, 2007 the presence of tufts of diactines. The tufts of diactines in P. perlucida are also regularly spaced ( Klautau 2013, personal communication), as we observed in P. s o l a n g e a e sp. nov. Nevertheless, these species can be easily distinguished, mainly by the sizes of their spicule categories, by the layer of tangential triactines that is present only in P. perlucida , and by the composition of the atrial skeleton, which is formed by triactines and tetractines in P. solangeae sp. nov. but is formed exclusively by tetractines in P. perlucida . Moreover, in P. perlucida , a second subatrial skeleton is occasionally present.
Paraleucilla sphaerica Lanna et al., 2009 View in CoL also resembles P. solangeae View in CoL sp. nov. Here are the main differences between these species: the presence of microdiactines [94.5 – 177.5 ± 5.8 – 315/ 9.0 ± 1.2 µm] in the cortical and choanosomal skeleton of P. sphaerica View in CoL , the presence of diactines [160 – 498.0 ± 103.7 – 850/ 14.3 ± 2.8 µm] arranged in tufts in P. solangeae View in CoL sp. nov., and the abundance of subatrial tetractines in P. solangeae View in CoL sp. nov., which are comparatively rare in P. sphaerica View in CoL .
The tufts of diactines also represent a key difference between P. solangeae View in CoL sp. nov. and the two new species described here (see below).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paraleucilla solangeae
| Cavalcanti, Fernanda F., Menegola, Carla & Lanna, Emilio 2014 |
Paraleucilla sphaerica Lanna et al ., 2009
| Lanna et al. 2009 |