
Decimodrilus bulbosus, Dózsa-Farkas & Nagy & Felföldi & Hong, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5094.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:86B2DB64-22B5-424A-81DC-91790DC4B209 |
|
DOI |
https://doi.org/10.5281/zenodo.6301528 |
|
persistent identifier |
https://treatment.plazi.org/id/C44F87E9-FFCC-FFB8-FF6E-06E7FB39F8D2 |
|
treatment provided by |
Plazi |
|
scientific name |
Decimodrilus bulbosus |
| status |
sp. nov. |
Decimodrilus bulbosus sp. n.
( Figures 2A View FIGURE 2 , 3–4 View FIGURE 3 View FIGURE 4 )
Type material. Holotype: NIBRIV0000886161, slide No. 2876, adult, stained whole mounted specimen [last 7 segments, 0.76 mm used for DNA analysis (No. 1395, ID number 911b)]. Type locality: Jeoksangsan , Jeoksangmyeon, Muju-gun, Jeollabuk-do, Korea, soil and litter layers of Pinus densiflora and mixed forests, N 35˚56′58.63″, E 127˚40′26.90″, 387 m asl, 06.11.2019 . Paratypes: In total 4 specimens, same data as for the holotype: NIBRIV0000886163, slide No. 2875, NIBRIV0000886162, slide No. 2877, P.137.1–137.2 slide No. 2874, 2878, adult, stained, whole mounted specimens .
Further material examined. No additional specimens were found at the study sites.
Etymology. It was named after the bulbiform ampulla of the spermatheca (bulbosus = bulbiform, Latin).
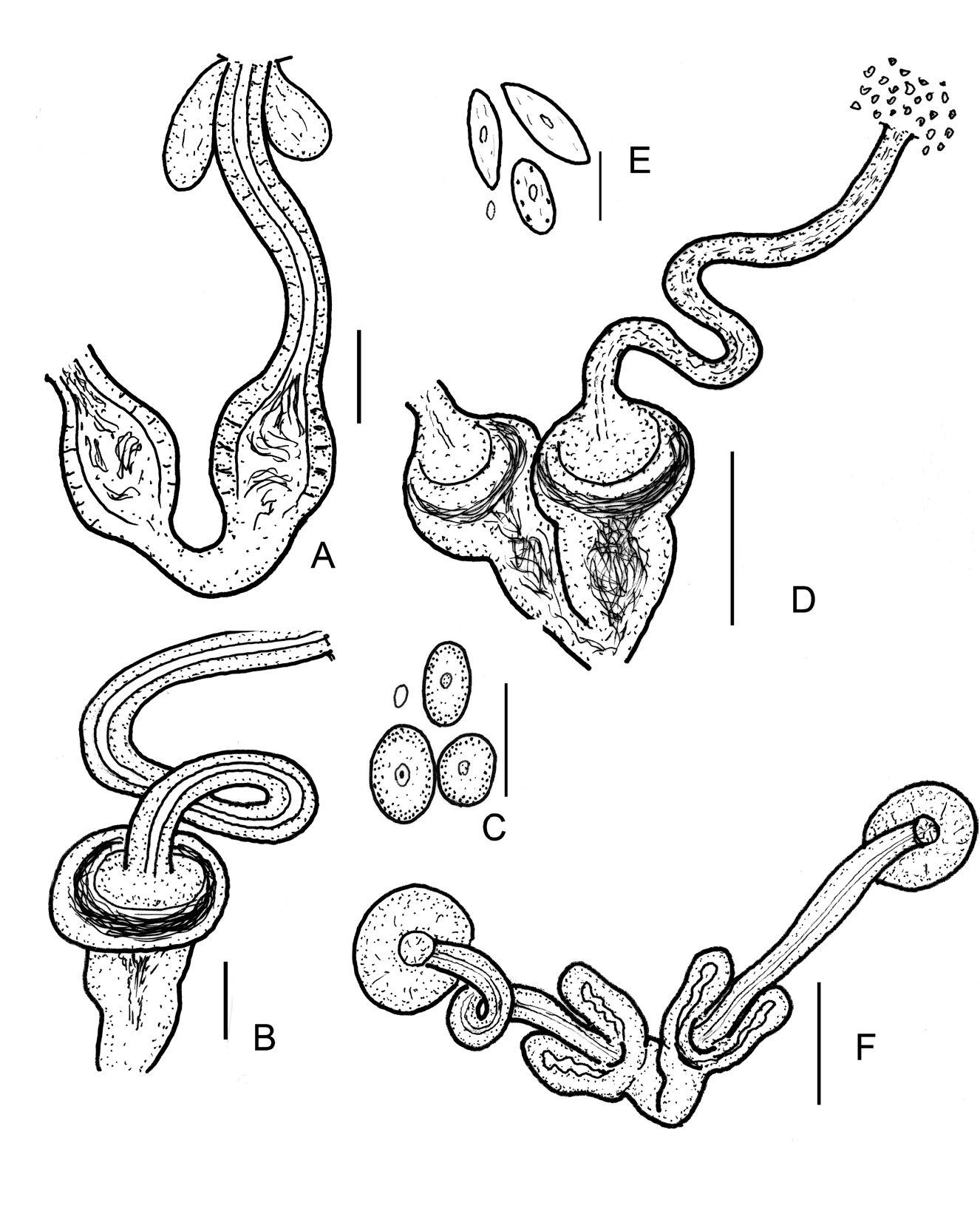
Diagnosis. The new species can be recognized by the following combination of characters: (1) 5.4–8.9 mm long ( in vivo), segments 33–39; (2) chaetae maximum 5–6 per bundle, straight or slightly bent in a bundle arranged in asymmetric fan without nodulus; (3) five pairs of prelitellar nephridia, 6/7–10/11, anteseptale consisting of funnel only; (4) dorsal lobes of pharyngeal glands usually free, connected in IV in some specimens, ventral lobes in IV–VI, small in IV, large in VI, one pair of secondary glands in VI; (5) dorsal blood vessel origin in XII, blood colourless; (6) intestinal diverticulum in IX–X; (7) sperm funnel cylindrical, 200–370 μm long, about 3–4 times longer than wide ( in vivo); (8) seminal vesicle absent; (9) spermathecal ectal duct contractile, 100–250 μm long and 20–30 μm wide ( in vivo), with one or two ectal glands and a relative large ampulla (diameter 50–75 μm), ampullae united proximally and connected jointly with oesophagus; (10) one mature egg at a time.
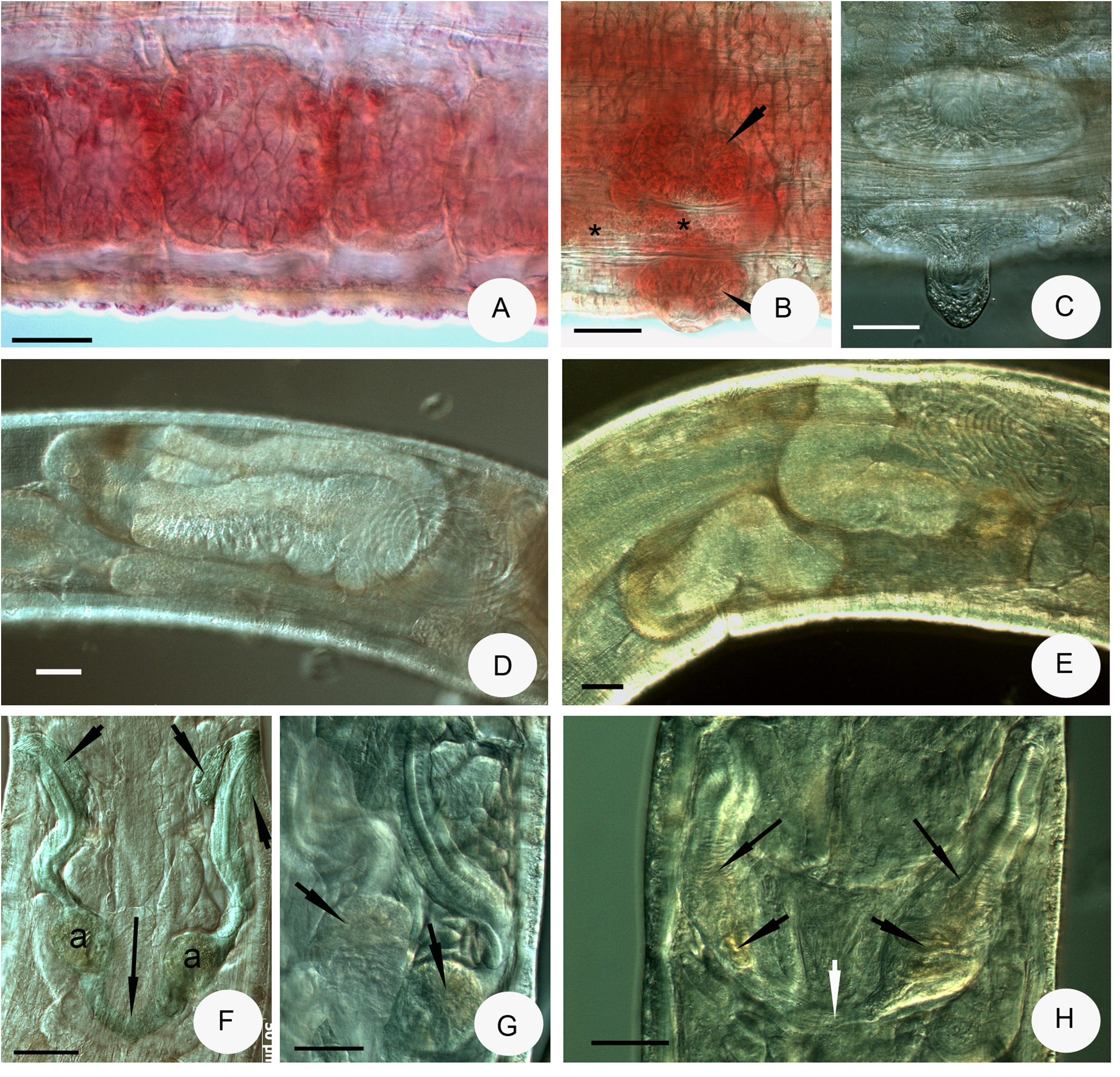
Description. Small worms. Holotype 8.9 mm long, 260 μm wide at VIII and 300 μm at clitellum in vivo ( 4.1 mm long, 250 μm wide at VIII and 265 μm at clitellum, fixed), 37 segments. Length of paratypes 5.4–8.9 mm, width 200–300 μm at VIII and 250–300 μm at clitellum in vivo, length of fixed specimens 3.4–4.9 mm, width 190–280 μm at VIII and 200–265 μm at clitellum, segments 33–39. Chaetal formula 3,4,5–3,4,(2): 5,6,4–4,5,3,(2). Chaetae straight or slightly bent without nodulus, in a bundle arranged in asymmetric fan, those towards dorsal and ventral midlines of body gradually smaller, e.g. 45, 35, 28, 23, 20 μm long in a ventral bundle of segment VI. Maximum length 38–45 μm, alike in preclitellar region and posterior segments, width about 2–3 μm, chaetae absent in XII. Epidermal glands conspicuous in vivo, in 4 transverse rows ( Fig. 3E View FIGURE 3 ). Body wall 20–28 μm thick, cuticle less than 1 μm. Clitellum extending over XII–½ XIII, girdle-shaped, gland cells irregularly scattered; larger hyaline gland cells are surrounded by smaller granular gland cells ( Fig. 3C View FIGURE 3 ); clitellum absent midventrally between the bursal slits and before them, posteriorly the structure same kind as dorsally. Head pore 0/I, a longitudinal rhombus-shaped slit ( Fig. 3D View FIGURE 3 ).
Brain ( Figs 3A,B View FIGURE 3 ) about 130 μm long in vivo and 95 μm when fixed, 1.5–2 times longer than wide, anteriorly slightly convex or concave, posteriorly truncated. Oesophageal appendages absent. Postpharyngeal bulbs well developed. All pairs of pharyngeal glands are free or united in IV dorsally, all with ventral lobes (in IV small, in VI large), and one small pair of secondary glands in VI, at times hardly visible. Intestinal widening (intestinal diverticula?) in IX–X ( Figs 3G–I View FIGURE 3 ), 130–170 μm wide in vivo (100–153 μm when fixed), about 50–70 μm wider than intestine before and after intestinal widening. In D. bulbosus the name of diverticula probably not correct, but the widening of the intestine is probably functionally identical with the well visible diverticula in D. diverticulatus . Chloragocytes about 13–16 μm high. Dorsal blood vessel from XII, blood colourless; the anterior bifurcation in peristomium. Five pairs of preclitellar nephridia from 6/7 to 10/11; anteseptale consisting of funnel only, efferent duct arises antero-ventrally, posteriorly the efferent duct arises posteriorly. Coelomocytes oval ( Fig. 3F View FIGURE 3 ), mucocytes with fine granulation and well visible nucleoli, about 28–37 μm long in vivo. Midgut pars tumida in XIX–XXIV occupying 3–5 segments ( Fig. 4A View FIGURE 4 ). Seminal vesicle absent. Sperm funnels ( Figs 3G View FIGURE 3 , 4D–E View FIGURE 4 ) cylindrical, 200–370 μm long, 3–4 times longer than wide in vivo (120–275 μm long and 2.5–4 times longer than wide in fixed specimens). The collar about as wide as funnel body or slightly narrower. Diameter of sperm ducts 6–7 μm. Spermatozoa 90–100 μm long, heads 35–40 μm in vivo (80–85 and 15–37 μm, fixed, respectively). Male glandular bulb ( Fig. 4B–C View FIGURE 4 ) compact, well developed, 130–170 μm long, 73–82 μm wide and 50–67 μm high in vivo (70–100 × 50–60 × 42–58 μm respectively, fixed). Spermathecae ( Figs 2A View FIGURE 2 , 4F–H View FIGURE 4 ) jointly attached to the oesophagus; ectal ducts contractile and 100–250 μm long and 20–30 μm wide in vivo (40–110 μm long, and 23–25 μm wide, fixed), canal well visible, 7–8 μm wide, distally widening at ampulla to 17–22 μm width in vivo, with one or two hyaline ectal glands ( Fig. 4F View FIGURE 4 ). Ectal glands often difficult to see because of large amounts of coelomocytes and, in the fixed specimens, because of the weak staining intensity. Ampullae onion-shaped, lumen with globular sperm masses ( Fig. 4F–H View FIGURE 4 ). Ampullae united proximally via short ental ducts and connected jointly with the oesophagus. One mature egg at a time.
Distribution and habitat. Only known from type locality (Site 2): Jeoksangsan, Jeoksang-myeon, Muju-gun, Jeollabuk-do, Korea, soil and litter layers of Pinus densiflora and mixed forests, N 35˚56′58.63″, E 127˚40′26.90″, 387 m asl.
Differential diagnosis. This new species belongs to Decimodrilus on account of the following characters: intestinal widening present in IX–X, the shape of brain, the number of preclitellar nephridia (5), the type of chaetae, secondary pharyngeal glands in VI, seminal vesicle absent, and the spermathecae connect jointly with the oesophagus. Currently, this species is the largest in the genus, and the principal differences are in spermathaecal ampulla: in D. diverticulatus Dózsa-Farkas, Nagy, Felföldi & Hong, 2019 the ampulla has two diverticula-like protrusions, in D. globulatus Dózsa-Farkas, Nagy, Felföldi & Hong, 2019 2–3 regular sperm rolls are in the onionshaped ampulla, whereas in the new species the large onion-shaped ampulla is filled only with a sperm mass not in rolls. The clitellum is entirely open ventrally in D. diverticulatus , but closed posteriorly in the new species and in D. globulatus . The sperm funnel is larger in D. bulbosus sp. n. (200–370 μm long and 2.8–4.8 times longer than wide but 70–130 μm long and 1.7–3.5 times longer than wide in D. globulatus and 90–140 μm long and 2–3 times longer than wide in D. diverticulatus in vivo). Similarly, the male copulatory organs of D. bulbosus sp. n. are larger than in the other species (130–170 μm long, vs. 115–140 μm in D. globulatus and 80–90 μm in D. diverticulatus in vivo). The maximum number of chaetae in a bundle is 5–6, but four in D. diverticulatus (five only in the case of one specimen) and five in D. globulatus (in one specimen, 6 in a single bundle). Furthermore, the intestinal widening in IX–X is stronger developed in D. diverticulatus than in the new species and in D. globulatus .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |