
Marcusenius altisambesi, Kramer & Skelton & Bank & Wink, 2007
|
publication ID |
https://doi.org/10.1080/00222930701250987 |
|
persistent identifier |
https://treatment.plazi.org/id/C512B407-FFD9-A040-E7D0-1368FB95DA93 |
|
treatment provided by |
Carolina |
|
scientific name |
Marcusenius altisambesi |
| status |
sp. nov. |
Marcusenius altisambesi View in CoL , sp. n.
( Figure 1 K View Figure 1 )
Gnathonemus okavangensis Pappenheim 1907 View in CoL . ‘‘Appeared as a form of Gnathonemus macrolepidotus Peters View in CoL from the Okavango R., Damaraland, Africa; regarded as infrasubspecific and not available’’ ( Eschmeyer 2006). ‘‘Nomen dubium’’ according to Gosse (1984) and Seegers (1996, p. 73).
Gnathonemus macrolepidotus: Gilchrist and Thompson 1913, pp. 330–331 View in CoL .
Type specimens. Holotype: SAIAB 79135 (specimen L39isi), Namibia: Caprivi Strip: Lisikili on Upper Zambezi River . Paratypes: SAIAB 79136 (6), SAIAB 79137 (3), ZSM 35086 (5) , ZSM 35085 (2) , ZSM 35097 (1) , ZSM 35082 (2).
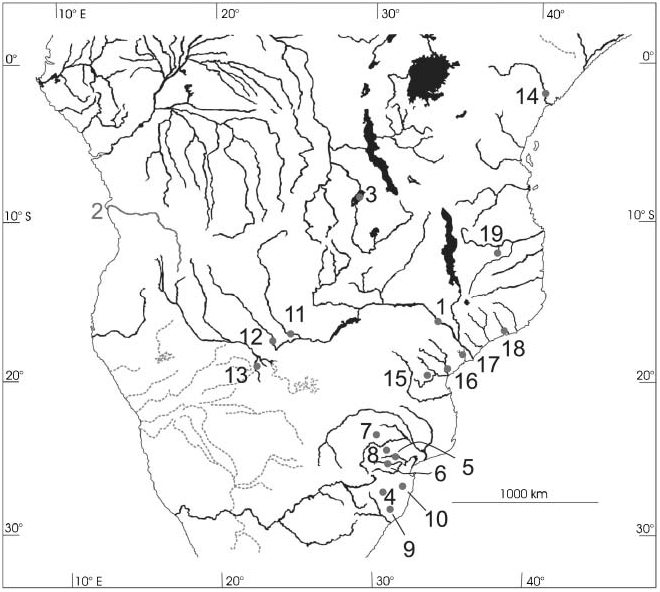
Type locality. Upper Zambezi in East Caprivi, specifically Upper Zambezi River comprised between Lisikili and Kalimbeza (or Kalambesa , 17 u 339S, 24 u 299E to 17 u 32927.30S, 24 u 31926.20E; 22–26 km straight line downstream from Katima Mulilo ; no. 11, Figure 4 View Figure 4 ) .
The first record of M. macrolepidotus for the Upper Zambezi is that of Gilchrist and Thompson (1917, p. 562; then termed Gnathonemus macrolepidotus ), specifying Lialui, Barotseland as the origin. For a description, the authors refer to Gilchrist and Thompson (1913, p. 330), a description of South African specimens. The presence of G. macrolepidotus in the Upper Zambezi System was confirmed by Jubb (1958). We disagree with Gilchrist and Thompson’s view (1917, p. 562) that their description of a South African specimen of Gnathonemus macrolepidotus (in Gilchrist and Thompson 1913) closely matches that of G. macrolepidotus from the Upper Zambezi.
Another possible synonym for our new species would be G. okavangensis if it were available. ‘‘ Gnathonemus okavangensis ’’ (apostrophes by Pappenheim) is not available because, first, Pappenheim (1907) stated that he would perhaps suggest that name if certain conditions were met (such as more material, better conservation status, sufficient differentiation). Second, there is neither a formal description nor a fish body deposited at a Museum with an accession number. Third, the origin ‘‘Okavango- Fluss (Damaraland, D.-S.-W.-Afrika)’’ of the single specimen sent to the Berlin Museum by Oberleutnant Volkmann in 1904 is somewhat mysterious since Damaraland is centred on Windhoek (confirmed by consulting a German map from the time in question), a dry region that is far off the Namibian, or any other, part of the Okavango River. In our opinion ‘‘ G. okavangensis ’’ should be removed from the list of nominal species for M. macrolepidotus ; it has not even been relegated simply because it never existed.
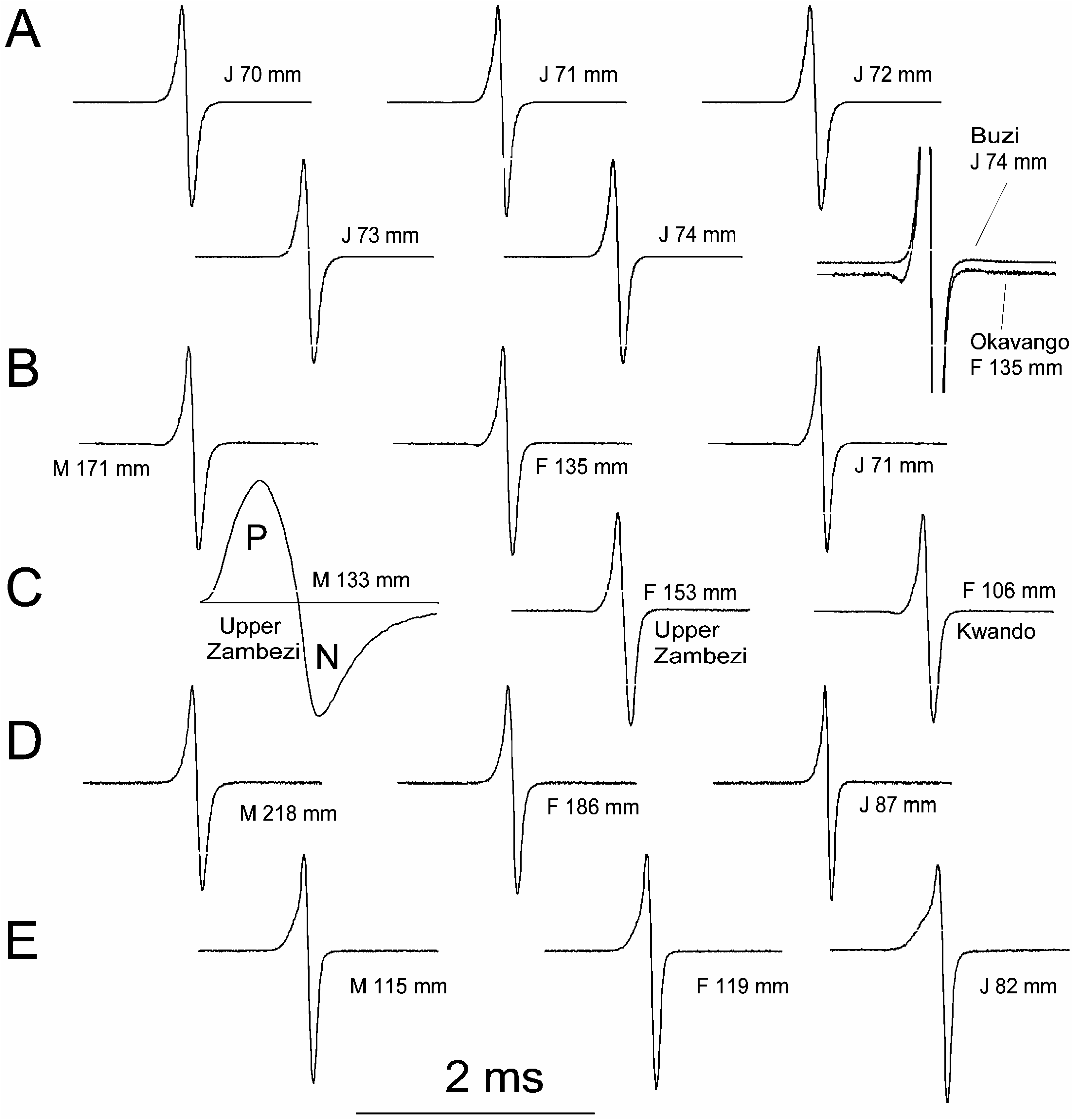
Diagnosis. A median SPc of 12 (12–14; Okavango,12–12), median SLS of 54 (49–60; Okavango, 51 – 57 – 57), median 29 (26–30) anal fin rays, median 23 (20–25; Okavango, 20 – 23 – 26) dorsal fin rays, deep-bodied with mean BD 27.5 % (20.1–32.8% of SL; Okavango, 21.8% – 28.1% – 32.5%), other mensural body proportions summarized in Table XI; EOD with weak head-negative pre-potential in juveniles and females, sexual dimorphism in EOD waveform present in ‘‘summer’’ in the form of greatly increased EOD duration in sexually mature males of greater than about 12.5 cm SL, but dimorphism not confirmed for Okavango specimens .
Description. Head with terminal mouth well in front of eye, mental lobe on lower jaw protruding beyond upper jaw. Head and body dorsolaterally compressed. Dorsal fin situated about two thirds of standard length from snout, obliquely orientated, anteriorly higher and posteriorly lower, distal margin sometimes only slightly crescentic with anterior two or three rays longer than posterior rays, number of rays 20 ( N 51), 21 ( N 56), 22 ( N 530), 23 ( N 535), 24 ( N 516), 25 ( N 51) [ Okavango specimens, similar distribution]; anal fin opposite dorsal fin with distinctly more anterior origin, obliquely orientated, anteriorly lower and posteriorly higher, anterior rays longer than posterior ones, especially in males where they also appear stronger and often darkened, distal margin crescentic (in males only posterior to rounded, elongated anterior part of fin), number of rays 26 ( N 54), 27 ( N 57), 28 ( N 532), 29 ( N 527), 30 ( N 519) [ Okavango specimens, similar distribution but mode at 29 rather than 28]. Scales cycloid with reticulate striae, scales extending anteriorly to operculum and pectoral fins (beyond pelvics). Scales on caudal peduncle circumference, 12 ( N 583), 13 ( N 53), 14 ( N 53) [ Okavango, 12 in all specimens]. Caudal peduncle relatively deep, subcylindrical entire length, usually 19% (16–22%) in SL (Table XI). Electric organ discharge biphasic with weak pre-potential in juveniles and females ( Figure 3 View Figure 3 ). In sexually mature males greater than about 12.5 cm SL, sexual dimorphism in the form of greatly increased EOD duration in ‘‘summer’’ (up to 611), not confirmed for specimens from Okavango. Males approaching sexual maturity develop a kink in the base of the anal fin (e.g. Figure 1K View Figure 1 ) which is absent in juveniles and females where the anal fin base is straight. Colour in life: beige-grey, head yellow-gold with greenish hue, paired fins yellow, dark brown blotches except on head, fewer on belly, purple shimmer depending on the angle of light incidence. Okavango specimens: similar to Upper Zambezi fish but darker, brownish grey with many distinct dark-brown blotches, less on belly, paired fins beige rather than yellow.
Colour in preservation. Specimens from the Upper Zambezi: medium brown with darker, irregular blotches on the sides. Narrow dark zone on the back not seen from the side. Belly same colour as body sides. Darkness of head reduced by opaque mormyrid skin (carrying electroreceptor organs, absent on the body sides). Specimens from the Okavango: similar to those from the Upper Zambezi, however, clearly darker, including the fins. Increasingly darker from belly to back.
Ecology. The Upper Zambezi River at Caprivi ( Namibia) is a free ranging, major system with regular flooding of a vast savannah plain at high altitude (. 900 m above sea level) that is covered by fine sand. The Zambezi in this region is characterized by major side channels giving rise to secondary and tertiary arms with a weaker current. It is a permanent river although the side channels of higher order, and especially peripheral pans, may be temporary or seasonal. At low water level, steep river banks may rise 10 m high. The tributary of the Upper Zambezi, the Kwando River , lacks high banks at least in Caprivi, and is more like the major Okavango River in its West except for its much smaller size, and by also dying in the savannah (however, without forming a vast inland delta). The Kwando River is occasionally flooded by the Upper Zambezi via Chobe , Linyanti , and Lake Liambezi. Lake Liambezi dried up completely in 1985 (except for brief periods of flooding by the Zambezi, e.g. in October 2001 when it was 28% full). During the day, bulldogs are found in rooted reed areas in main channels as well as inside floating reed mats in side channels like at Lisikili. These mats may be 2 m deep, and fish leave cover only at night. Huge numbers of small juveniles may be found in grassy pans on the periphery of major river arms or side channels in the dry season. The Okavango ecology is similar, but differing by an extensive system of lake-like lagoons, where water chestnut, water lilies and dense papyrus and reed beds are common .
Distribution. Occurs in the Upper Zambezi River system in Namibia’s Caprivi Strip, including the lower Kwando River (including its terminal parts named Linyanti River). Because these rivers are international borders, this system is bordered by Zambia in its north, Botswana south, Zambia and Zimbabwe east. The species’ downstream (southeastern) limit is assumed to be the nearby Victoria Falls ( Zimbabwe and Zambia), as with many other Upper Zambezi fish species. Northern limits in Zambia and Angola are not yet explored. The species also occurs in the western Okavango System ( Botswana, Namibia, Angola). Limits still further to the west, e.g. the Cunene River, have not been sufficiently explored.
Relationships. Marcusenius altisambesi is considered closest to M. macrolepidotus based largely on the confusion of the identity of both species, and the occurrence in the same river, the Zambezi. Marcusenius altisambesi is distinguished most easily by the circumferential caudal peduncle scale count of 12 vs. 16, whereas medians 23 vs. 22 for dorsal ray counts, and 29 vs. 28 for anal ray counts, are less distinct. In M. altisambesi when compared to M. macrolepidotus the anterior body part is relatively longer and body depth higher, as shown by high PDL, PAL, and BD measurements; by contrast, head length HL is shorter. The mental lobe is longer in M. altisambesi than in M. macrolepidotus . In female and juvenile EOD small head-negative prepotential is present in M. altisambesi but not M. macrolepidotus (from Buzi River; Figure 3 View Figure 3 ); Upper Zambezi bulldog males have a sexually dimorphic EOD of long duration when in the breeding condition. Another close relationship is G. moeruensis of which Boulenger (1915) states that it is ‘‘très voisin’’ to G. angolensis . However, with nA 5 33, both G. angolensis and G. moeruensis are beyond the maximum observed in our large sample of M. altisambesi ( nAmax 530). G. moeruensis also differs by HL which is greater than the 90th percentile observed in M. altisambesi (Table XI).
Etymology. Marcusenius altisambesi refers to the Upper Zambezi River, that is, the section of the Zambezi that ends at Victoria Falls (with ‘‘sambesi’’ as a noun in apposition).

| PAL |
Herbarium Mediterraneum Panormitanum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Marcusenius altisambesi
| Kramer, Bernd, Skelton, Paul, Bank, Herman Van Der & Wink, Michael 2007 |
Gnathonemus macrolepidotus
| : Gilchrist and Thompson 1913: 330 - 331 |
Gnathonemus okavangensis
| Pappenheim 1907 |