
Ronzotherium filholi ( Osborn, 1900 )
|
publication ID |
https://doi.org/10.5852/ejt.2021.753.1389 |
|
publication LSID |
lsid:zoobank.org:pub:8009DD3B-53B0-45C9-921E-58D04C9C0B48 |
|
DOI |
https://doi.org/10.5281/zenodo.4959526 |
|
persistent identifier |
https://treatment.plazi.org/id/C53FFD4D-D71E-FF84-FDAF-DD751945A543 |
|
treatment provided by |
Felipe |
|
scientific name |
Ronzotherium filholi ( Osborn, 1900 ) |
| status |
|
Ronzotherium filholi ( Osborn, 1900)
Figs 11–14 View Fig View Fig View Fig View Fig
Aceratherium filholi Osborn, 1900: 240–243 , figs 7, 8a.
Badactherium latidens Croizet, 1841: 79 (nomen nudum).
Rhinoceros brivatensis Bravard, 1843: 408–410 (nomen oblitum).
Rhinoceros incisivus Blainville, 1846 : pl. XII (Ongulogrades, ‘Auvergne’) (misidentification).
Rhinoceros minutus Thomas, 1867: 239 (misidentification).
Rhinoceros tetradactylus Filhol, 1877: 126 (misidentification).
Rhinoceros lemanensis Lydekker, 1886: 153 (from Caylux) (misidentification).
Praeaceratherium minus Koch, 1911: 377–379 , 385–387 (misidentification).
Paracaenopus kochi Kretzoi, 1940: 92 .
Ronzotherium filholi elongatum Heissig, 1969: 46–55 , figs 16–17, 18a–c, 19 (from Villebramar) (misidentification).
Ronzotherium velaunum – Aymard 1856: 235. — Boada-Saña et al. 2007: 6.
Rhinoceros brivatensis – Aymard 1856: 235.
Badactherium latidens – Landesque 1888: 21, 27.
Rhinoceros latidens – Landesque 1888: 27.
Aceratherium lemanense – Pavlov 1892: 184, pl. V, fig. 7 (from Quercy).
Ronzotherium filholi – Deninger 1903: 95. — Wood 1929: 2 (= “ Praeaceratherium minus ” = Paracenopus). — Lavocat 1951: 116–118 (from Bournoncle). — Brunet & Guth 1968: 573–575, pl. I. — Heissig 1969: 38. — Brunet 1970: 2535; 1979: 105–152, 159–161, figs 8, 9a, c, e, 10a, 11–14, 16b, pls IX–XIV, XVIa, XIXm–n, XX–XXV. — Santafé Llopis 1978: 44. — Antoine 2002: 32. — Becker 2003: 231, pl. IIh (from Bressaucourt); 2009: 493–495, fig. 4g (from Bressaucourt).
Praeaceratherium filholi – Abel 1910: 18–20, 44–45.
Acerotherium filholi – Roman 1910: 1559 (from Quercy and Puylaurens); 1912a: 5, 27, 45, 51–53, fig. 16a (from Quercy, Villebramar and Puylaurens).
Praeaceratherium filholi – Koch 1911: 377–379, 385–386. — Wood 1927: 232 /72.
Acerotherium lemanense – Roman 1912a: 60–61 (from Montans).
Aceratherium filholi – Stehlin 1914: 185 (from Bressaucourt).
Paracaenopus filholi – Breuning 1924: 7, 17–20, figs?6, 7.
? Aceratherium filnoli [sic] – Crusafont Pairó 1967: 116.
Ronzotherium filholi filholi – Heissig 1969: 39–46, figs 12–15, 25c–d, 26a–b.
Ronzotherium kochi – Heissig 1969: 36–37. — Adrover et al. 1983: 126. — Codrea & Şuraru 1989: 322. — Guérin 1989: 4. — Uhlig 1999a: 477–479. — Codrea 2000: 38–42, fig. 8.
Epiaceratherium ? kochi – Brunet 1979: 158.
Allacerops kochi – Russell et al. 1982: 58.
“ Ronzotherium ” kochi – Radulescu & Samson 1989: 302.
Epiaceratherium sp. – Becker 2009 (= Ronzotherium kochi ).
Non Ronzotherium filholi – Lavocat 1951: 116,pl.19 fig. 3,pl.26 fig. 1(from Vendèze)(misidentification).
Non Ronzotherium filholi – Brunet 1979: 105, 134 (from Pernes, Kleinblauen and Bumbach) (misidentification).
Non Ronzotherium filholi – Becker 2003: 230–233, pl. IIa–f (from Kleinblauen and Bumbach) (misidentification).
Non Ronzotherium filholi – Becker 2009: 493–495, fig. 4h–l (from Kleinblauen) (misidentification).
Historical diagnosis
(From Osborn 1900): “Large upper premolars, simple, unlike molars, with incompletely formed crests; upper molars with internal cingulum and strong protoconule [= paracone] fold, small antecrochet, no crochet; depression in posterior face of metaloph of third molar; third and fourth lower premolars with depressed and incomplete posterior crests. Measurements: P2–M3=224.”
However, this diagnosis could refer to several species of Ronzotherium since these characters are mostly synapomorphies of the genus. Therefore, we emend the diagnosis based on the type specimens from the Phosphorites du Quercy. Other emended diagnoses were provided by Heissig (1969) and Brunet (1979), but they were not only based on the type material, but also on referred material from other localities. We emend here the diagnosis based on our phylogenetic analysis.
Emended diagnosis
The coronoid process of the mandible is rather weak. The upper premolars are large, simple, nonmolariform, with incompletely formed protoloph and metaloph, and labial cingulum always present; P2 molariform, protocone and hypocone usually fused on P3–4, strong, simple and continuous lingual cingulum, usually without ridges; crista sometimes present on P3; metaloph of P2–4 discontinuous; upper molars with strong and continuous lingual cingulum except under the hypocone of M1, almost no labial cingulum, small antecrochet, no crochet, and a posterior groove on the ectometaloph of M3; lower cheek teeth with strong and continuous labial cingulum and lingual cingulum in the opening of the posterior valley; d/p1 usually present and two-rooted, the paraconid of p2 is developed; the magnum facet of the McII is straight; high proximal articulation of the fibula with the tibia; the expansion of the calcaneus facet is wide and low on the astragalus; proximal border of the anterior side of the MtIII straight and intermediate reliefs of the metapodials low and smooth.
It differs from R. velaunum by the deep median constriction of the distal humeral articulation and from R. elongatum by its close frontoparietal crests, its straight occipital crest and its poorly developed processus posttympanicus and its constricted metaloph on P3–4 (hypocone not connected to the ectoloph).
It further differs from R. elongatum and R. romani by its sharp angle at the anterior tip of the zygomatic process and the higher posterior side of the scaphoid compared to its anterior side.
Type material
Holotype
FRANCE • maxilla fragment with right and left cheek teeth rows with P2–M3; Quercy Phosphorites (southwestern France); MNHN.F.QU7232 .
Paratypes
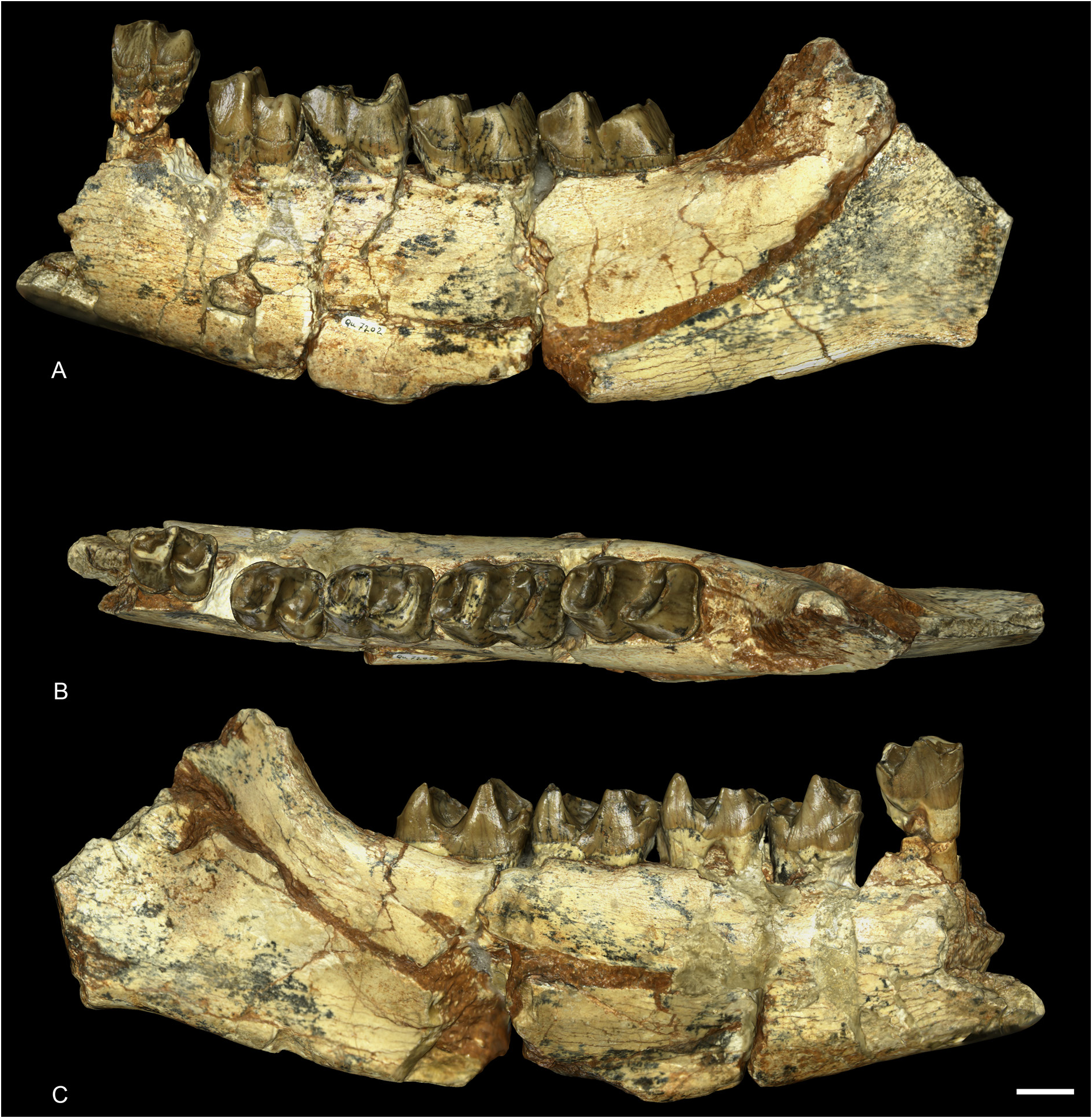
FRANCE • 1 left mandible fragment; Quercy; MNHN.F.QU7202 • 1 right mandible fragment; Quercy; MNHN.F.QU7201 .
Osborn (1900) designated a left mandible fragment ( MNHN.F.QU7202) also from Quercy as “cotype”, which was followed by Heissig (1969), who also added its right counterpart ( MNHN.F.QU7201) from the same individual. These two hemimandibles should be regarded as paratypes. The upper and lower anterior dentition are unknown.
Additional material
Old collections from Quercy are preserved in almost every large European institution, including, but not limited to the MNHN, TLM or NMB, but are problematic because the exact age and locality are unknown. The specimens examined from these collections that we mention in the text are:
FRANCE – Quercy • 1 right maxillary fragment with P1-2; MNHN.F.QU16445 • 1 left hemimandible with m1-3; MNHN.F.QU17193 • 1 right scaphoid; NMB-QV-275 • 1 right lunate; NMB-QE-440 • 1 left pyramidal; NMB-QE-433 • 1 left magnum; NMB-QE-472 • 1 left cuboid; NMB-QE-362 . – Bournoncle- Saint-Pierre • 1 astragalus; MNHN.LIM7 .
ROMANIA – Cluj-Napoca • 1 right maxilla with P2–M3; MBT 1509 .
GERMANY – Espenhain • 1 left radius; BSPG-2008-I-44 . – Möhren 4 • 1 left D4; BSPG-1966 - XXXIII-47 • 1 left MtIV; BSPG- 1971-V . – Möhren 7 • 1 left P1; BSPG-1969-XXIV-151 • 1 left P3; BSPG-1969-XXIV-150 • 1 right p3/4; BSPG-1969-XXIV-71 • 1 fragment of left lower molar; BSPG- 1969-XXIV-152 • 1 right distal ulna; BSPG-1969-XXIV • 1 right proximal McIII; BSPG-1969-XXIV • 1 fragmentary astragalus; BSPG-1969-XXIV-183 • 1 right MtII; BSPG-1969-XXIV-73 • 1 left MtIII; BSPG-1969-XXIV-156 . – Möhren 11 • 1 right calcaneum; BSPG-1971-V-11 .
Type horizon and locality
Unknown horizon and locality in the Phosphorites du Quercy.
Stratigraphical distribution
Possibly restricted to the early Oligocene.
Geographical distribution
France: Phosphorites du Quercy, Bournoncle Saint-Pierre, Villebramar, Penchenat (= Moulinet?), Puylaurens. Germany: Möhren 4, 7/16, 19, 20, Burgmagerbein 8, Ronheim 1, Grafenmühle 6. Romania: Cluj-Napoca. Spain: Montalbán. Switzerland: Bressaucourt.
Description
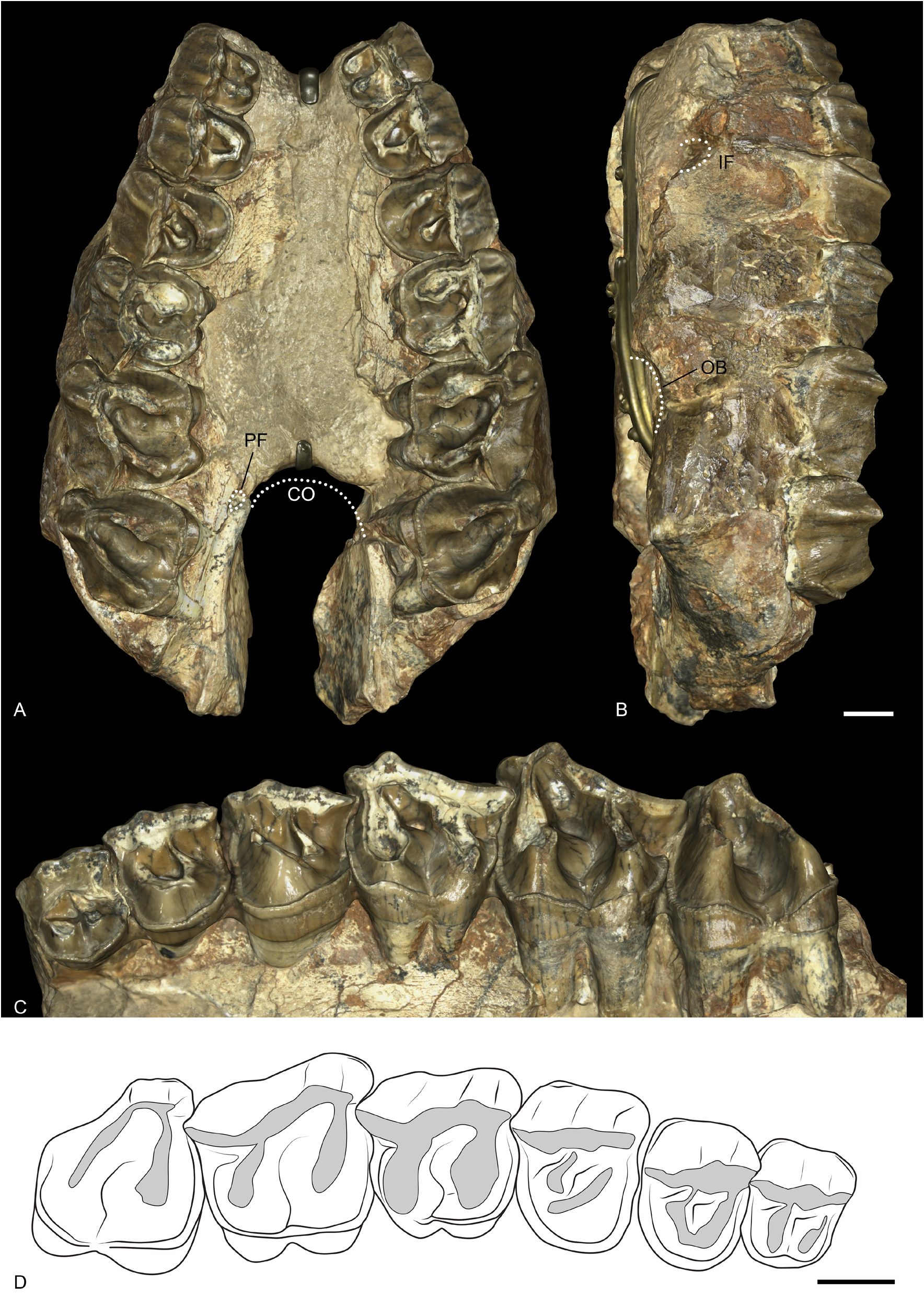
MAXILLA. The right and left maxillae of the holotype MNHN.F.QU7232 are well preserved, and bear P2–M3 on both sides ( Fig. 11 View Fig ). The anterior border of the choanae opens approximately at the level of M2 and the palatine foramen is at the level of the anterior border of M3. The infraorbital foramen (still preserved via the infraorbital canal) is located above the anterior border of P4. The anterior border of the orbit is between M2 and M1. The zygomatic arches are broken but the anterior border was above M2 and was high above the teeth neck. The retromolar space behind M3 is short.
MANDIBLES. The paratype hemimandibles MNHN.F.QU7202 and MNHN.F.QU7201 are incomplete, the symphysis and the two rami are not preserved ( Fig. 12 View Fig ). The foramen mentale was anterior to p3. The base of the corpus mandibulae is straight and the lingual groove is present, though extremely shallow, and barely visible. The foramen mandibulare is located below the teeth neck line. Because of the fragmentary condition of the specimen, no other characters can be observed. From another mandible from Quercy (MNHN.F.QU17193), we can observe that the posterior border of the mandible and the foramen mentale were both located at the level of p2, lingually and labially.
UPPER DENTITION. The cheek teeth have no cement and the crown is low ( Fig. 11 View Fig ). The LP3–4/LM1–2 ratio is equal to 0.51, i.e., the premolar row is long compared to the molar row.
The first premolar is not preserved on the holotype MNHN.F.QU7232. Only one P1 was found among the numerous isolated teeth of Ronzotherium from Quercy in the MNHN collection, on a maxilla fragment with P2 (MNHN.F.QU16445). It has three roots, two labial and a lingual one. The paracone is the largest cusp, and the paracone and metacone folds are strong. The protocone is extremely weak, and fuses with the strong and continuous lingual cingulum. The protocone connects lingually to the hypocone by a small bridge. The protoloph is very weak and does not fully connect to the paracone. The metaloph is complete and connects the well-developed hypocone to the metacone. The parastyle is weak. The anterolingual cingulum is present. The labial cingulum is strong under the parastyle and the metacone but absent under the paracone.
All upper premolars (P2–4) on the holotype have a very strong and continuous lingual cingulum, which extends anteriorly and posteriorly. The labial cingulum is only present under the parastyle and metastyle, and completely absent under the paracone and metacone. The paracone fold is rather strong and the metacone fold is weak. There is no constriction of the protocone. They have no crista, crochet or antecrochet and the postfossette is narrow. They all bear three roots.
The protocone and hypocone of P2 are equal and connected by a low lingual bridge. The protoloph is weak and directed towards the parastyle, not the paracone, and does not fully connect to the ectoloph. The metaloph is continuous and postero-lingually directed.
On P3, the hypocone is very weak and very poorly differentiated from the protocone by a shallow lingual groove. The protoloph is straight, connected to the parastyle and well developed. The metaloph is thinner, transverse and S-shaped.
The protocone and hypocone of P4 are completely fused, and the protoloph is L-shaped. The metaloph is very weak and it is completely separated from the protocone/hypocone. It is S-shaped, short and connects to the ectoloph between the paracone and the metacone.
Upper molars have four roots. The lingual cingulum is strong and continuous, except under the hypocone of M1, where it is completely fainted. The labial cingulum is almost completely absent except for a few traces either under the parastyle or the metastyle. The paracone fold is strong and the metacone fold is absent. There is a broad and weak mesostyle on the ectoloph of M1. The crochet, crista and medifossette are completely absent and there is no protocone constriction. The posterior part of the ectoloph is straight.
The M1 is square. The antecrochet is broad and distinguished by a postero-lingual groove on the protoloph. The postfossette is very short and shallow. The metaloph and protoloph are transverse. The posterior cingulum is high and continuous.
The M2 differs from M1 by its larger size, the more oblique lophs, a shorter metaloph, and the metacone more lingual. There is no lingual groove of the protocone. The mesostyle is very weak and disappears at the base of the crown.
The M3 is quadrangular but bears no metacone. The metaloph and ectoloph are fused into an ectometaloph. The protocone is not constricted and the protoloph is transverse. The posterior groove on the ectometaloph is present.
LOWER DENTITION. The p1, p2 and anterior dentition are unknown from the paratypes MNHN.F.QU7202 and MNHN.F.QU7201, and from the other mandibles from Quercy ( Fig. 12 View Fig ).
Other lower cheek teeth (p3–m3) are double-rooted, low-crowned, and have no cement. The labial cingulum is strong and almost completely continuous, it only vanishes under the ectolophid groove and it is very weak overall on m1. The lingual cingulum is present at the opening of the anterior and posterior valleys. The ectolophid groove is developed until the neck. In occlusal view, the trigonid is very angular and forms a right dihedron while the talonid is rounded. The metaconid of p3 bears a weak anterior crest that is almost joining the anterior branch of the paralophid. There are no vertical rugosities on p3. The talonid of p3–4 is poorly developed and the entoconid is almost completely absent. The hypolophid is very low and the posterior valley is U-shaped in lingual view. The anterior valley opens much higher above the neck than the posterior one. The metaconid of premolars is very large and slightly constricted. The anterior branch of the paralophid is long on molars and premolars. The entoconid of molars is strongly developed and slightly constricted.
POSTCRANIALS. The postcranial remains from the Quercy collection can only be hardly associated with the cranial remains for several reasons. First, almost all specimens belong to ‘old’ collections, i.e., the exact localities were not specified, and specimens were mixed together and could belong to several loci. Furthermore, the Quercy localities range in age from the early Eocene to the early Miocene, and thus cannot be precisely dated. Therefore, only a few well-preserved postcranial remains are tentatively attributed to? R. filholi and described here.
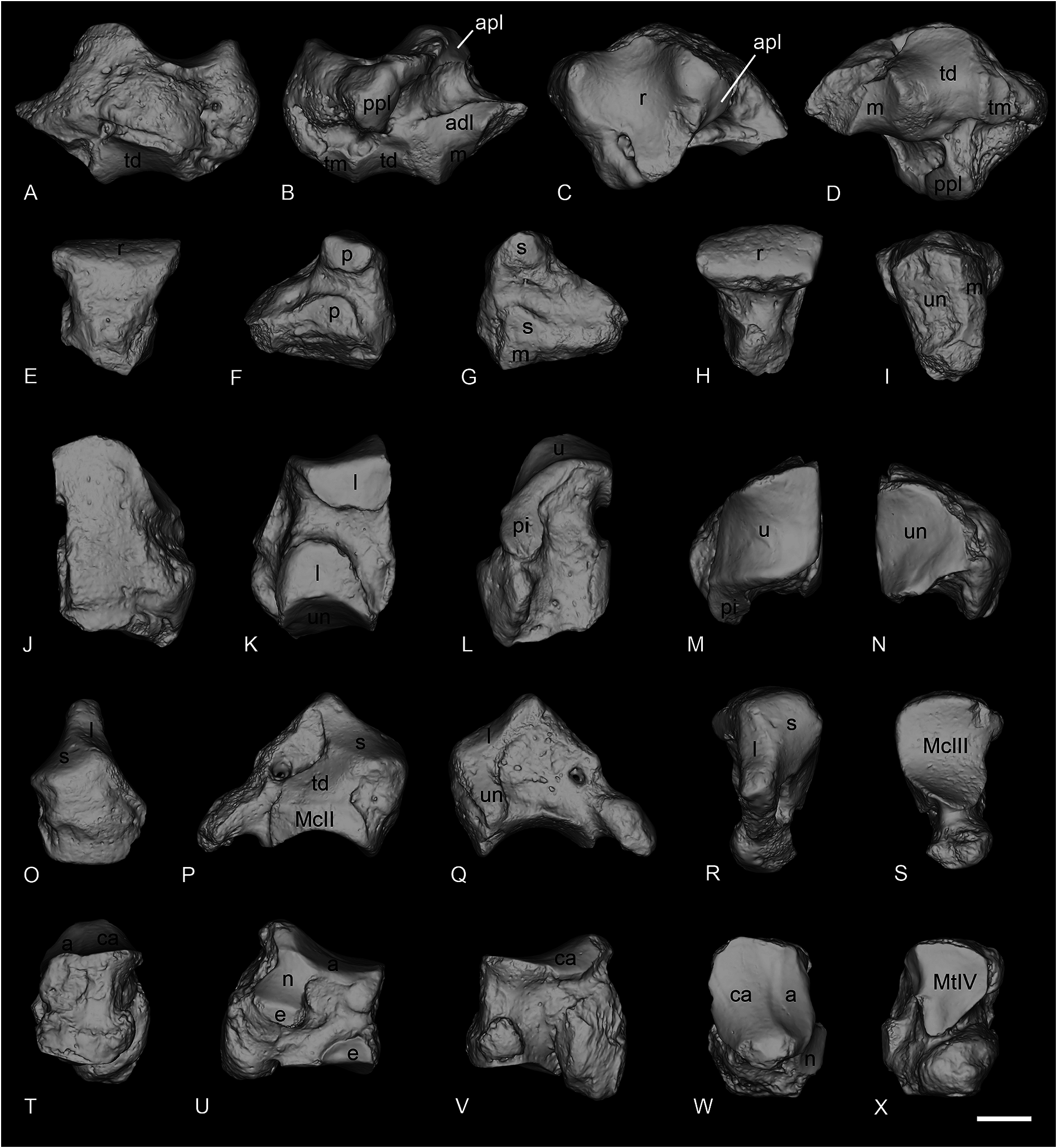
SCAPHOID. The scaphoid NMB-QV-275 is very well preserved, except for the distal part of the anterior apophysis, which is partly broken ( Fig. 13A–D View Fig ). The posterior height is slightly reduced compared to the anterior. The proximal articulation for the radius is large, and very concave anteroposteriorly. It is lozenge-shaped in proximal view, and very developed laterally. Below and anterior to this proximal facet is the thin and elongated anteroproximal facet for the lunate, which is completely fused to the posteroproximal one. The anteroproximal one is horizontal while the posterior is oblique. The anterodistal facet for the lunate is separated from the proximal ones by a wide groove. This facet is long and low, but hardly distinguishable from the distal magnum facet just below. This distal facet for the magnum is very concave in lateral view and separated from the large medio-distal facet for the trapezoid by a high ridge. The latter is also very concave in lateral view, but very convex mesio-laterally, and bears a large extension on the medial side. The trapezium facet is not reduced and separated from the trapezoid facet by a ridge. It is quite flat and oval-shaped.
LUNATE. The lunate NMB-QE-440 is very poorly preserved and the posterior part is broken ( Fig. 13E–I View Fig ). In anterior view, the distal border is very acute. Three facets are visible in medial view, two small ones are for the scaphoid, while the most distal one, for the magnum, is thin and elongated until the posterior border. On the lateral side, the two facets for the pyramidal are separated by a deep groove. The distal facet is larger than the proximal one. In distal view, the unciform facet is large, almost rectangular and anteroposteriorly concave.
PYRAMIDAL. The pyramidal NMB-QE-433 is perfectly preserved ( Fig. 13J–N View Fig ). The proximal articulation for the ulna is very large, concave anteroposteriorly and convex transversally. The postero-proximal facet for the pisiform is long and drop-shaped. On the medial side, there are two large facets for the lunate, separated by a deep groove. The distal one is symmetrical and slightly curved towards the posterior side. In distal view, the facet for the unciform is triangular and concave anteroposteriorly.
MAGNUM. The magnum NMB-QE-472 is well preserved and complete ( Fig. 13O–S View Fig ). The anterior side is pentagonal, and the proximal apophysis is very high. In anterior view, the anterior border of the scaphoid facet is slightly concave while the distal border is almost completely straight. On the lateral side, the lunate and unciform facets are fused and form a unique L-shaped facet, occupying all the anterolateral part of the proximal apophysis. Distally, it contacts the distal facet for the McIII. This latter is very concave anteroposteriorly. The posterior tuberosity is very short and curved. In medial view, the distal McII facet and the medial facet for the trapezoid are almost in the same plane; they are only distinguished by a very low ridge, and there is no indentation separating them. The McII facet is flat. The trapezoid facet is large and widely connected to the proximal magnum facet.
CUBOID. The cuboid NMB-QE-362 is perfectly preserved ( Fig. 13T–X View Fig ). In proximal view, the posterior apophysis is almost not visible. The proximal articulation is oval-shaped, and the two surfaces for the astragalus and calcaneus are very poorly distinguished. It is very concave anteroposteriorly and very high at the posterior end. The proximal border of the anterior side is oblique, the distal one is slightly convex while the medial and lateral borders are irregular and are crossed by median grooves. The lateral one is much wider than the medial one and isolates the posterior apophysis. On the medial side, the anterodistal facet for the ectocuneiform is large and anteroposteriorly elongated. The postero-proximal facet for the navicular is very large, concave and bears a thin anterior extension below the proximal articulation. The small and rectangular posterior surface for the ectocuneiform is located almost perpendicular to the postero-distal border of this navicular surface. The distal articulation for the MtIV is a triangular-shaped lozenge and is deeper than wide.
Remark
Ronzotherium filholi is also known from other localities, notably in Villebramar and Bournoncle-Saint- Pierre, where specimens are rather well-preserved. It is found at several localities of South-Germany (e.g., Möhren, Burgmagerbein or Ronheim; Uhlig 1999a), but only by scarce remains. We also consider R. kochi from Cluj-Napoca ( Fig. 14 View Fig ) as a junior synonym of R. filholi . All the material from Villebramar has already been fully described by Brunet (1979) so it will not be described again here. The locality of Villebramar provided by far the broadest sample for R. filholi , including a complete skull, several hemimandibles and numerous postcranial remains.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Rhinocerotoidea |
|
Family |
|
|
Genus |
Ronzotherium filholi ( Osborn, 1900 )
| Tissier, Jérémy, Antoine, Pierre-Olivier & Becker, Damien 2021 |
Ronzotherium filholi
| Becker D. 2009: 493 |
Ronzotherium filholi
| Becker D. 2003: 230 |
Ronzotherium
| Radulescu C. & Samson P. 1989: 302 |
Allacerops kochi
| Russell D. E. & Hartenberger J. - L. & Pomerol C. & Sen, S. & Schmidt-Kittler N. & Vianey-Liaud M. 1982: 58 |
Epiaceratherium
| Brunet M. 1979: 158 |
Ronzotherium filholi
| Brunet M. 1979: 105 |
Ronzotherium filholi elongatum
| Heissig K. 1969: 55 |
Ronzotherium filholi filholi
| Heissig K. 1969: 39 |
Ronzotherium kochi
| Codrea V. 2000: 38 |
| Uhlig U. 1999: 477 |
| Codrea V. & Suraru N. 1989: 322 |
| Guerin C. 1989: 4 |
| Adrover R. & Feist M. & Ginsburg L. & Guerin C. & Hugueney M. & Moissenet E. 1983: 126 |
| Heissig K. 1969: 36 |
Aceratherium filnoli
| Crusafont Pairo M. 1967: 116 |
Ronzotherium filholi
| Lavocat R. 1951: 116 |
Paracaenopus kochi
| Kretzoi M. 1940: 92 |
Paracaenopus filholi
| Breuning S. von 1924: 7 |
Aceratherium filholi
| Stehlin H. G. 1914: 185 |
Acerotherium lemanense
| Roman F. 1912: 60 |
Praeaceratherium minus
| Koch A. 1911: 379 |
Praeaceratherium filholi
| Wood H. E. 1927: 232 |
| Koch A. 1911: 377 |
Praeaceratherium filholi
| Abel O. 1910: 18 |
Acerotherium filholi
| Roman F. 1910: 1559 |
Ronzotherium filholi
| Becker D. 2003: 231 |
| Antoine P. - O. 2002: 32 |
| Brunet M. 1979: 105 |
| Santafe Llopis J. 1978: 44 |
| Brunet M. 1970: 2535 |
| Heissig K. 1969: 38 |
| Brunet M. & Guth C. 1968: 573 |
| Lavocat R. 1951: 116 |
| Wood H. E. 1929: 2 |
| Deninger K. 1903: 95 |
Aceratherium filholi
| Osborn H. F. 1900: 243 |
Aceratherium lemanense
| Pavlov M. 1892: 184 |
latidens
| Landesque 1888: 21 |
Rhinoceros latidens
| Landesque 1888: 27 |
Rhinoceros lemanensis
| Lydekker R. 1886: 153 |
Rhinoceros tetradactylus
| Filhol H. 1877: 126 |
Rhinoceros minutus
| Thomas P. 1867: 239 |
Ronzotherium velaunum
| Boada-Sana A. & Hervet S. & Antoine P. - O. 2007: 6 |
| Aymard A. 1856: 235 |
Rhinoceros brivatensis
| Aymard A. 1856: 235 |
Rhinoceros brivatensis
| Bravard A. 1843: 410 |
latidens
| Croizet J. B. 1841: 79 |