
Echinoplana celerrima Haswell, 1907
|
publication ID |
https://doi.org/10.5281/zenodo.275104 |
|
DOI |
https://doi.org/10.5281/zenodo.6213981 |
|
persistent identifier |
https://treatment.plazi.org/id/C6628794-3338-FF85-FF42-466F8BEDFC57 |
|
treatment provided by |
Plazi |
|
scientific name |
Echinoplana celerrima Haswell, 1907 |
| status |
|
Echinoplana celerrima Haswell, 1907 View in CoL
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Distribution in Tunisia: Several specimens were collected from the lagoon of Tunis: a natural lagoon located between the city of Tunis and the Gulf of Tunis, ( 36°48 ' 24.40" N; 10°14 ' 54.53" E).
Other localities: Australia: Haswell (1907), Johnston & Lee (2008). New Zealand: Holleman (2007). Italy: Galleni (1979), Ballarin & Galleni (1984).
Material: Of 10 specimens sectioned sagittally, two were fully sexually mature. The material examined was deposited in the Museo Nacional de Ciencias Naturales de Madrid ( Spain).
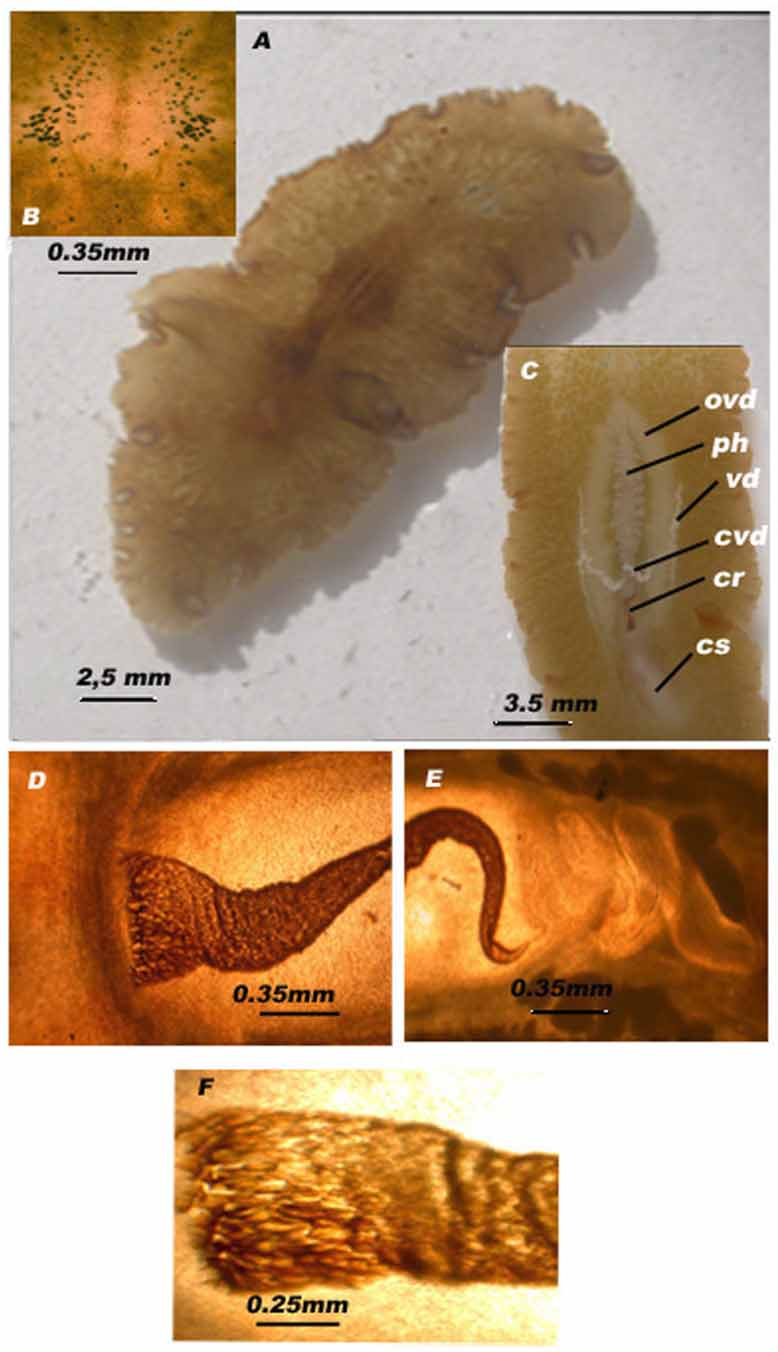
Description: Oval shaped body, brownish colour. A median dark band is present on the dorsal surface ( Fig. 1 View FIGURE 1 A). The length of the living specimens collected varies between 10 and 37 mm and the width from 4 to 18 mm. Tentacles typical to the family are lacking in this species but anterior to the brain there is a small protrusion visible especially on animals fixed with Bouin Hollande. The cerebral and tentacular eyes ( 35–65 in number) form two elongated paramedian bands. The tentacular eyes are wider than the cerebral ones and are the most posterior ( Fig. 1 View FIGURE 1 B). The ventral surface is transparent and colourless ( Fig. 1 View FIGURE 1 C). The body wall is composed of the basement membrane and three muscular layers (longitudinal-circular-longitudinal). The epidermis is nucleated and provided with rhabdites.
The observation of the pale ventral surface revealed the ruffled plicate pharynx located in the second quarter of the body. The mouth lies at the anterior part of the second third of the body and opens posterior to the centre of the pharyngeal cavity.
Male copulatory apparatus: The testes are generally arranged ventrally. The vasa deferentia extend latero-caudally forming two paramedian lines parallel to the pharynx, and join together behind the pharynx, forming a common vas deferens ( Fig. 1 View FIGURE 1 C). External observation of the ventral surface, as well as histological sections, show that the vasa deferentia are in close contact with the ventral wall of the body.
The copulatory organ is seen by transparency on both surfaces ( Fig. 1 View FIGURE 1 C). The male copulatory organ consists of a seminal vesicle, an interpolated prostatic vesicle and a distal male atrium housing the cirrus ( Fig. 1 View FIGURE 1 D, E). The complex is surrounded by inner circular and outer longitudinal muscle layers.
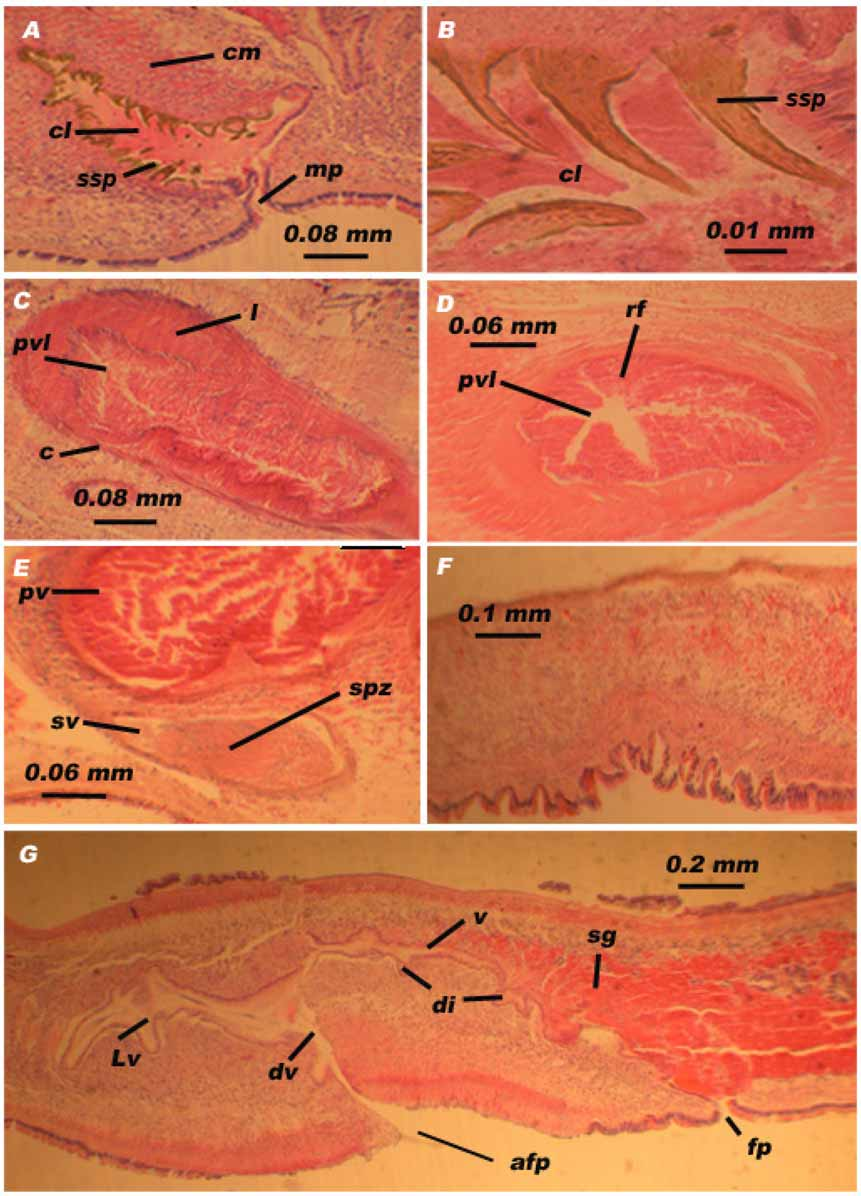
The seminal vesicle is covered with a thin muscular wall ( Fig. 2 View FIGURE 2 E) and is 4 to 5 times smaller than the prostatic vesicle. The seminal vesicle is usually filled with sperm cells ( Fig. 1 View FIGURE 1 C), and is connected, through a narrow tube, to the proximal end of the prostatic vesicle.
The prostatic vesicle is of an interpolated type and is characterized by a very thick muscular wall consisting of an external layer of longitudinal fibres and a well-developed internal layer of circular fibers ( Fig. 2 View FIGURE 2 C, D). The internal glandular epithelium has radially directed folds ( Fig. 2 View FIGURE 2 D). Our observations agree with the description of Galleni (1978) and Prudhoe (1982) but differ from the description given by Haswell (1907) who reported a horizontal orientation of the epithelial folds within the prostatic vesicle. The ejaculatory duct, surrounded by a muscular wall, runs from the distal part of the prostatic vesicle via the prostato-penial duct into the proximal part of the cirrus ( Fig. 3 View FIGURE 3 ).
c r: cirrus, c s: corrugated surface, c v d: common vas deferens, o v: oviduct, p h: pharynx, v d: vasa deferentia.
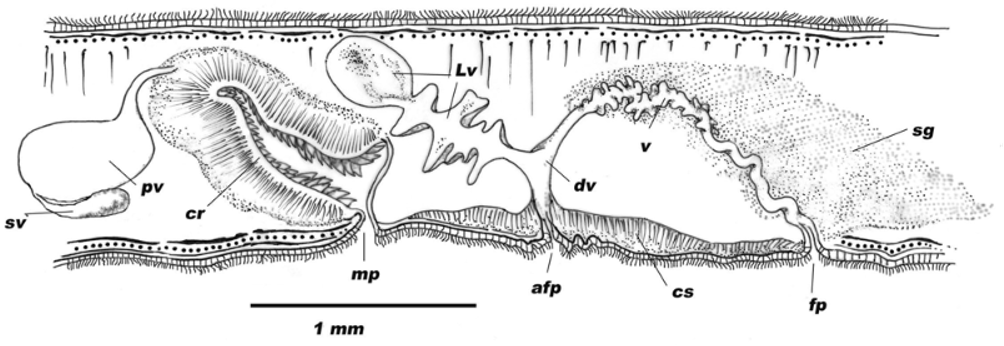
a f p: accessory female pore, c: circular muscle, c l: cirrus lumen, c m: cirrus muscle, d i: diverticle, d v: ductus vaginalis, f p: female pore, l: longitudinal muscle, L v: Lang’s vesicle, mp: male pore, p v: prostatic vesicle, p v l: prostatic vesicle lumen, r f: radial folds, s g: shell glands, s p z: sperm, s sp: sclerotized spine, s v: seminal vesicle, v: vagina.
The cirrus is caudo-frontally orientated and its inner lining is covered with sclerotized spines ( Fig. 1 View FIGURE 1 D, E). The size of the spines decreases proximally ( Figs.2 View FIGURE 2 A, B; 3). The largest spines reach up to 150 µm. The cirrus is normally retracted or invaginated in the male antrum but some specimens appear with an everted cirrus ( Fig. 1 View FIGURE 1 F).
Female copulatory apparatus: The ovaries lie more or less dorsally. In mature individuals, the female gonopore is located about 2 mm behind the male gonopore. The female gonopore leads into a long narrow vagina, which is divisible into three compartments: the vagina externa, the vagina media, and the vagina interna. The vagina externa is lined by a secretory epithelium. The vagina media, known as the ‘‘shell chamber’’, is oblique and frontally oriented. This section is characterized by abundant shell glands. The last section of the vagina is known as vagina interna. Near the proximal end, the vagina interna receives the common oviduct. In the majority of specimens, the vagina interna continues into a terminal portion with a prolongation which was described by Haswell (1907) as follows: “This unsymmetrical anterior prolongation of the vagina does not have a specially developed muscular layer, so that it cannot be looked upon as a bursa copulatrix”. During our histological study of the copulatory apparatus, two specimens presented a female apparatus with a relatively large Lang’s vesicle ( Fig. 2 View FIGURE 2 G and Fig. 3 View FIGURE 3 ). This elongate vesicle (about 1 mm) appears just where the vagina interna bends dorso-ventrally. It runs posteriorly and it is provided with large bilateral diverticles orientated caudo-ventrally ( Fig. 3 View FIGURE 3 ). Its epithelium is glandular, similar to that of the vagina and oviducts; it is surrounded by a thin, single-layered circular muscular wall. Sperm cells were seen inside the Lang’s vesicle and also the diverticles.
The Lang’s vesicle receives at the distal end the vagina interna and a relatively narrow accessory canal, the vaginal duct. This duct leads to the exterior through a third genital aperture ( Fig. 3 View FIGURE 3 ). The pore of the vaginal duct is oval shaped, transversally orientated and located between the female and male pore. Some mature specimens without a vaginal duct show an epidermal invagination which could be considered as the beginning of the development of the duct.
In living specimens as well as in fixed ones, a corrugated surface was observed on the ventral surface in front of the female pore. Histological sections allowed us to clearly see the structure of this surface, which appears in the form of small folds of various sizes ( Fig. 2 View FIGURE 2 F, G). The widest folds are located in the middle, in accordance with Holleman (2007) but contradicting the description by Galleni (1978), who mentions that the largest are those closest to the female pore. These structures are formed by a deformation of the ventral wall, which is covered by a short glandular epithelium which, in sections, shows a comb-shaped pattern ( Fig. 2 View FIGURE 2 F).
Remarks: Echinoplana celerrima Haswell, 1907 is the only species in the genus. The first description of this species was made by Haswell (1907) based on material sampled at Port Jackson, Sydney, Australia. In his revision of the Polycladida, Bock (1913) placed the genus Echinoplana in the family Planoceridae , based on the disposition of the eyes. Marcus & Marcus (1966) retained the species in the family Planoceridae but placed it in the subfamily Gnesiocerotinae Marcus & Marcus 1966. Galleni (1978) reported E. celerrima for the first time for the Mediterranean Sea, whereas Prudhoe (1982) once again described this species from Australia and raised the subfamily Gnesiocerotinae to family rank. Faubel (1983) accepted the taxonomic position within the family Gnesiocerotidae , characterised by the presence of an interpolated prostatic vesicle with ridged or folded interior glandular lining.
It appears that the majority of the sectioned individuals described in previous studies were not fully sexually mature. This assumption is based on the fact that the female apparatus described by Haswell (1907), Galleni (1978) and Prudhoe (1982), shows an incompletely developed Lang’s vesicle with the vaginal duct absent.
The study of Tunisian specimens revealed new morphological characteristics that may shed new light on the phylogenetic relationships. According to our results, E. celerrima is furnished with a well-developed Lang’s vesicle and a ductus vaginalis. In acotylean species, the ductus vaginalis opens either into the vagina externa or onto the exterior by a separate aperture located posteriorly to the female genital aperture (cf. Faubel 1983, Prudhoe, 1985).
The presence of a Lang’s vesicle and a ductus vaginalis demonstrated that E. celerrima can present three mature stages during its sexual differentiation process. First stage: the individual develops a female apparatus with a vagina provided with diverticles, the Lang’s vesicle is lacking and both oviducts become uteri ready to receive oocytes. Second stage: the Lang’s vesicle begins to develop. Third stage: The Lang’s vesicle develops completely and communicates with the exterior by a proper duct: the ductus vaginalis.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |