
Pristimantis jamescameroni, Kok, 2013
|
publication ID |
https://doi.org/ 10.5852/ejt.2013.60 |
|
publication LSID |
lsid:zoobank.org:pub:67D17AEB-B622-4184-9265-D3D2D6DA500A |
|
DOI |
https://doi.org/10.5281/zenodo.3843695 |
|
persistent identifier |
https://treatment.plazi.org/id/67EB6C36-0F31-4587-9805-3136BEF4E010 |
|
taxon LSID |
lsid:zoobank.org:act:67EB6C36-0F31-4587-9805-3136BEF4E010 |
|
treatment provided by |
Carolina |
|
scientific name |
Pristimantis jamescameroni |
| status |
sp. nov. |
Pristimantis jamescameroni sp. nov.
Figs 2-5 View Fig View Fig View Fig View Fig , Table 1 View Table 1
urn:lsid:zoobank.org:act:67EB6C36-0F31-4587-9805-3136BEF4E010
Eleutherodactylinae series b (in part) – Gorzula & Señaris 1999: 55 + plate 46.
Eleutherodactylus View in CoL sp. B – McDiarmid & Donnell, 2005: 514 [table 18A.1], 524.
Pristimantis sp. – Hedges et al. 2008: 179.
Pristimantis sp. “Aprada” – Kok et al. 2012: Suppl. Information: 13.
Definition and diagnosis
A small species of the genus Pristimantis currently not assigned to any species group, but morphologically most similar to species of the polyphyletic unistrigatus species group (sensu Hedges et al. 2008, see also discussion in Kok et al. 2011) mainly characterized in having Finger I shorter than II, Toe V longer than III, extending to the distal edge of the distal subarticular tubercle of Toe IV when toes are adpressed, and by the absence of cranial crests and the presence of vomerine teeth. The new species is characterized by the following unique combination of characters: (1) body small, male 22.8 mm SVL, females 26.3-27.5 mm (n=3); (2) dorsal skin shagreen, belly skin coarsely areolate; (3) tympanum absent (tympanic membrane not differentiated and tympanic annulus not visible externally); (4) tiny pharyngeal ostia present; (5) snout rounded to subovoid in dorsal view, rounded to slightly sloping in profile, canthus rostralis nearly straight in dorsal view, rounded in cross section, loreal region slightly concave, flaring slightly at upper lip; (6) upper eyelid shagreen with 1-2 distinctly enlarged tubercles on each eyelid; (7) choanae small, oval, dentigerous processes of vomers very small, barely visible, slightly oblique, ovoid to triangular, posterior and medial to choanae, each bearing 1-5 teeth; (8) absence of vocal slits in male; (9) tongue cordiform; (10) two large, mostly unpigmented nuptial pads on each thumb in male; (11) Finger I shorter than II; (12) fingers with lateral fringes, best developed preaxially on Fingers II-III; (13) ulnar tubercles absent or inconspicuous, when present low, not forming a distinct line; (14) tarsal tubercles absent, one inconspicuous calcar tubercle present; (15) inner metatarsal tubercle oval, about four times the size of the round, projecting outer metatarsal tubercle; (16) Toe V longer than III, extending to the distal edge of the distal tubercle on Toe IV when toes are adpressed; (17) toes with lateral fringes, best developed preaxially on Toes III-IV, webbing basal between Toes IV-V; (18) in life main dorsal background colouration is orange to orangish dark brown, either with irregular dark brown chevrons or a darker middorsal band, and/or a light middorsal stripe, ventral colouration is white with brown reticulation and an orange patch posteriorly; (19) in preservative dorsal background colouration is light to dark brown, either with irregular brown chevrons or a darker middorsal band, and/or a light middorsal stripe, ventral colouration is cream with brown reticulation.
Etymology
The specific epithet is a noun in the genitive case, honouring the Canadian film director, producer, environmentalist and explorer Mr. James F. Cameron in recognition of his efforts to alert the general public to environmental problems through pioneering high quality “blockbuster” movies and adventurous documentaries. James Cameron also encourages people to go vegan (a diet excluding animal products), one of the effective ways to reduce human environmental impacts such as global climate change, identified as a serious threat to tepuis ecosystems (see Nogué et al. 2009).
Material examined
Holotype
VENEZUELA: an adult male collected by Philippe J. R. Kok, 15 Jun. 2012 at 08h18, summit of Apradatepui , Bolívar state, 5°24’42”N, 62°27’00” W, 2570 m elevation, IRSNB 4160 (field number PK 3636).
GoogleMapsParatopotypes (n=4)
VENEZUELA: three adult females (IRSNB 4161-63, field numbers PK 3632, PK 3637, PK 3635) and one juvenile (IRSNB 4164, field number PK 3639), collected by Philippe J. R. Kok (except IRSNB 4161, collected by Brad Wilson) 14-15 Jun. 2012, all from the summit of Aprada-tepui between 2557- 2571 m elevation.
Description of the holotype
An adult male 22.8 mm SVL ( Figs 2 View Fig A-B, 3, 4D, H, see Table 1 View Table 1 for measurements), in very good condition, except a large scar under the right thigh where a piece of muscle was removed prior to preservation. Head slightly longer than wide (HW 94.7% of HL), wider than body; HW 39.0% of SVL; HL 41.2% of SVL; cranial crests absent. Snout longer than eye length (SL 117.1% of EL), rounded to subovoid in dorsal view, rounded to slightly sloping in profile; canthus rostralis nearly straight in dorsal view, rounded in cross section, loreal region slightly concave, flaring slightly at upper lip; eye-naris distance shorter than eye length (EN 71.4% of EL). Nares slightly protuberant, directed posterolaterally, visible in frontal and dorsal views. Widest upper eyelid width narrower than interorbital distance (upper eyelid width 82.7% of IO), upper eyelid shagreen with 1-2 enlarged tubercles on each eyelid. Tympanum absent (tympanic membrane not differentiated and tympanic annulus not visible externally); tiny pharyngeal ostia present. Supratympanic fold conspicuous in life, slightly arched, originating at posterior corner of eye, failing to reach shoulder; post-rictal tubercles evident. Choanae small, oval, not concealed by palatal shelf of maxillary arch; dentigerous processes of vomers very small, barely visible, slightly oblique, ovoid to triangular, posterior and medial to choanae, each bearing 3-4 teeth. Tongue cordiform, slightly longer than wide, rounded posteriorly, posterior half free. Vocal slits and vocal sac absent.
Dorsal skin shagreen, including on head; middorsal raphe detectable; no dermal folds or ridges visible on dorsal surface; flanks granular. Throat surface shagreen; upper chest shagreen, slightly “wrinkled”; weak thoracic fold; belly skin coarsely areolate; weak discoidal fold anterior to groin; posteroventral thigh and cloacal region coarsely areolate; cloacal sheath absent.
Hand length 27.2% of SVL. Finger I 80.0% of II. Relative length of fingers III> IV> II> I; adpressed Finger I fails to reach proximal edge of digital pad of Finger II; adpressed Finger IV reaches the intercalary cartilage of Finger III on the left side, the base of the disc of Finger III on the right side. Two large, not connected, non-spinous, mostly unpigmented (translucent when wet, white when dry) nuptial pads on each thumb, one extending along the preaxial surface of the thenar tubercle and invading most of it, the other one extending along the dorsal and the preaxial surface of the thumb. Lateral fringes on all fingers, best developed preaxially on Fingers II-III ( Fig. 3 View Fig ). Finger discs broadly expanded, elliptical, broader than long, circumferential groove conspicuous, distal edge of disc rounded; disc of Finger III 2.1 times wider that the distal end of the adjacent phalanx. Palmar tubercle large, poorly defined, not fully pigmented, deeply bifid; thenar tubercle large, protuberant, ovoid; supernumerary tubercles few, large (almost equal in size to the subarticular tubercles), slightly protuberant; subarticular tubercles large, round and protuberant, one each on FI and FII, two each on FIII and FIV. Ulnar tubercles few, inconspicuous, forming an ill-defined line; presence of a small antebrachial tubercle.
Hind limbs moderate in length, heels slightly overlap when held at right angles to sagittal plane; TIL 50.4% of SVL; FL 41.7% of SVL. Relative length of Toes IV> V> III> II> I; tip of Toe V extends to the distal edge of the distal subarticular tubercle on Toe IV; tip of Toe III extends to the distal edge of the penultimate subarticular tubercle on Toe IV on the left side, to the proximal half of the penultimate subarticular tubercle on Toe IV on the right side. Lateral fringes on all toes, best developed preaxially on Toes III-IV; webbing basal between Toe IV-V ( Fig. 3 View Fig ). Toe discs mostly equal in size to finger discs, WTD/WFD = 1; toe discs broadly expanded, elliptical, broader than long, circumferential groove conspicuous, distal edge of disc rounded. Inner metatarsal tubercle elongate, oval, about four times the size of the projecting, round outer metatarsal tubercle; subarticular tubercles round, large and protuberant; supernumerary plantar tubercles small, low and round, increasing in size distally. Single, inconspicuous and very small round calcar tubercle; no outer tarsal tubercles detectable; inner tarsal fold not detectable.
Colour of holotype in life (see Fig. 2 View Fig A-B)
Dorsal background colour orangish brown covered with numerous tiny white flecks, top of head slightly darker; inconspicuous darker (than adjacent dorsum) middorsal band outlined by ill-defined dark brown dorsolateral lines; irregular white band on the canthus rostralis; bluish white line on outer edge of upper eyelids; upper lips white; most post-rictal tubercles white; enlarged tubercles on eyelids light brown. Flanks orangish brown, with numerous tiny white flecks and some irregular white spots of variable sizes; groins, anterior thighs, and armpits bright orange. Arms and legs coloured as dorsal surfaces, with a few small irregular white spots on upper arms and on the distal part of tarsi. Throat, chest, and lower flanks white with brown anastomosed reticulation; belly and most of the undersurface of thighs bright orange; undersurface of distal thighs, shanks, and tarsi brown with a few small irregular white spots; posterior surface of thighs and cloacal area brown. Upper face of hands and feet orangish brown, except tip of Fingers I-II, which is bright orange, and tip of toes, which is whitish. Palms and soles brown; undersurface of Fingers I-III bright orange, including discs; undersurface of tip of Toes I-III bright orange including discs. Iris silver with dark brown venation and an ill-defined, broad horizontal brownish stripe; greenish silver spot on the posterior upper part of the iris.
Colour of holotype in preservative (see Fig. 4D, H View Fig )
After 13 months in 70% ethanol (July 2013), orangish brown faded to light brown. Orange and white faded to cream. Dorsal pattern turned generally more conspicuous. Brown reticulation on throat, chest, and lower flanks faded to light brown. Granules in the cloacal area, as well as subarticular and supernumerary tubercles became dark brown. Discs are greyish brown.
Sexual dimorphism and variation among paratypes
See Table 1 View Table 1 for measurements of the type series, and Figs 2 View Fig , 4 View Fig , 5 View Fig for intraspecific variation. Sexual dimorphism evident in size, with adult females being larger than the adult male (26.3-27.5 vs. 22.8 mm SVL), and by the presence of two nuptial pads in the male. A sexual dichromatism in belly colouration seems to occur, with the presence of an orange spot restricted to the middle area of the belly in living females, whereas the orange area covers the belly, the posterior part of the ventral surface, and the undersurface of thighs in a more continuous way in the living male (see Fig. 2B, D, F View Fig ). Hands and feet slightly longer in the male than in the females (HAND III 27.2% vs. 24.0-25.1% of SVL; FL 41.7% vs. 37.8-40.3% of SVL). No additional significant difference is detected in other size ratios. No significant variation in skin texture occurs among the preserved specimens, but in life IRSNB 4163 (adult female, Fig. 2C View Fig ) had a conspicuous dorsolateral fold extending from behind the eye to the end of the body; that fold totally disappeared in preservative. Eyelid tubercles are more difficult to detect in preserved specimens and range from 1 to 2 on each eyelid. Number of teeth on dentigerous processes of vomers varies from 2 to 6. Weak thoracic and discoidal folds detectable in IRSNB 4163 (female) and IRSNB 4164 (juvenile), not seen in the other paratypes. Tip of Toe V fails to reach the distal edge of the distal subarticular tubercle on Toe IV on the left side in IRSNB 4161 (female) only.
Colour pattern variable. In life, dorsal background colouration varies from bright orange to orangish dark brown, usually covered with tiny white flecks; dorsal pattern varies from irregular brown chevrons (e.g., in IRSNB 4161 and IRSNB 4162) to a slightly darker (than adjacent dorsum) middorsal band outlined by ill-defined dark brown dorsolateral lines (e.g., in IRSNB 4160) and/or a pale middorsal stripe (e.g., in IRSNB 4163); pattern on flanks varies from a few irregular oblique white (e.g., IRSNB 4163) or brown/white (e.g., IRSNB 4162) stripes to white spots (holotype); colour of ventral reticulation varies from brown to dark brown; inconspicuous transverse bands on arms and legs occur in two female paratypes (IRSNB 4162 and IRSNB 4163). Enlarged tubercles on eyelids are usually white (light brown in the holotype). The juvenile (IRSNB 4164, Fig. 5 View Fig ) has a complex and more marked dorsal pattern consisting of a greyish brown interorbital line preceded by a greyish brown circle and followed by a wide W-shaped marking outlined by greyish brown in the scapular region; the W-shaped marking is followed by a greyish brown arch, itself followed by two back to back C-shaped light grey lines. It also differs from adults in having an irregular greyish band on the canthus rostralis, the upper lip not completely white, granules in the loreal region, dark brown labial stripes, the supratympanic fold underlined by a dark brown band, brown transverse bands on the forearm, and reddish transverse bands on the legs. Iris varies from silver or greenish silver to bronze, with dark brown venation and an illdefined, broad horizontal brownish stripe.
Morphological comparisons with congeneric species
Available data suggest that tepui summit Pristimantis species have relatively restricted distributions in the Guinea Shield highlands; therefore, comparisons with congeners only focus on Pristimantis species known to occur in the Pantepui region (as defined by Kok 2013). Comparisons of external character states are based both on original descriptions and examination of museum specimens (see Appendix for material examined).
Twenty species of Pristimantis are currently reported from the Pantepui region: P. aureoventris Kok, Means & Bossuyt, 2011 , only known from the summit of Wei-Assipu-tepui and the upper slopes of Mount Roraima, Cuyuni-Mazaruni District, Guyana; P. abakapa Rojas-Runjaic, Salerno, Señaris & Pauly, 2013 , described from the summit of Abakapá-tepui, Bolívar state, Venezuela; P. auricarens ( Myers & Donnelly, 2008) , known only from Auyán-tepui, Bolívar state, Venezuela; P. avius (Myers & Donnelly, 1997) , reported from Pico Tamacuari, Amazonas state, Venezuela; P. cantitans (Myers & Donnelly, 1996) , known from Cerro Yaví and Cerro Yutajé, Amazonas state, Venezuela; P. dendrobatoides Means & Savage, 2007 , only known from the Wokomung massif, Potaro-Siparuni District, Guyana; P. guaiquinimensis ( Schlüter & Rödder, 2007) , described from Guaiquinima-tepui, Bolívar state, Venezuela; P. jester Means & Savage, 2007 , known from the Wokomung massif, Potaro-Siparuni District, Guyana, and the slopes of Maringma-tepui, Cuyuni-Mazaruni District, Guyana (Kok, pers. obs. 2007); P. marahuaka ( Fuentes-Ramos & Barrio-Amorós, 2004) , endemic to Cerro Marahuaka, Amazonas state, Venezuela; P. marmoratus (Boulenger, 1900) , reported as widespread in the Guiana Shield from eastern Venezuela to French Guiana and northern Brazil ( Frost 2013), but several species may hide under this name; P. memorans (Myers & Donnelly, 1997) , known only from the Sierra Tapirapecó, Amazonas state, Venezuela; P. muchimuk Barrio-Amorós, Mesa, Brewer-Carías & McDiarmid 2010 , apparently endemic to the summit of Churí-tepui, Bolívar state, Venezuela; P. pruinatus (Myers & Donnelly, 1996) , known only from Cerro Yaví, Amazonas state, Venezuela; P. pulvinatus (Rivero, 1968) , reported from the Gran Sabana region and Auyán-tepui, Bolívar state, Venezuela to western Guyana ( Frost 2013), but more than one species may hide under this name (see Kok et al. 2012); P. saltissimus Means & Savage, 2007 , known from the Wokomung massif, Potaro-Siparuni District, Guyana, and the slopes of Maringma-tepui, Cuyuni-Mazaruni District, Guyana (Kok pers. obs. 2007); P. sarisarinama Barrio- Amorós & Brewer-Carías, 2008, endemic to Sarisariñama-tepui, Bolívar state, Venezuela; P. vilarsi (Melin, 1941) , known from upper Amazonian Brazil, Colombia, Venezuela and Peru ( Frost 2013); P. yaviensis (Myers & Donnelly, 1996) , reported from Cerro Yaví and Cerro Yutajé, Amazonas state, Venezuela; P. yuruaniensis Rödder & Jungfer, 2008 , known with certainty only from the summit of Yuruaní-tepui, Bolívar state, Venezuela (possible occurrence on the neighbouring Kukenán-tepui, see Mägdefrau & Mägdefrau 1994; Rödder & Jungfer 2008; Kok et al. 2011); and P. zeuctotylus ( Lynch & Hoogmoed, 1977) , widespread in northeastern South America ( Frost 2013).
Pristimantis jamescameroni sp. nov. is most easily distinguished from P. abakapa , P. aureoventris , P. avius , P. cantitans , P. dendrobatoides , P. guaiquinimensis , P. marmoratus , P. memorans , P. pruinatus , P. pulvinatus , P. saltissimus , P. sarisarinama , P. vilarsi , P. yuruaniensis and P. zeuctotylus by lacking a differentiated tympanic membrane and an external tympanic annulus (at least one of these structures is detectable in the latter 15 species). It further differs from P. vilarsi and P. zeuctotylus in having Finger I <II (Finger I> II in P. vilarsi and P. zeuctotylus ).
Pristimantis jamescameroni sp. nov. is immediately distinguished from P. auricarens , P. jester , P. marahuaka , P. muchimuk , and P. yaviensis (the only known Pantepui “earless” Pristimantis ) in having conspicuous lateral fringes on fingers and toes (absent or limited to a weak keel in the latter 5 species).
Distribution and ecology
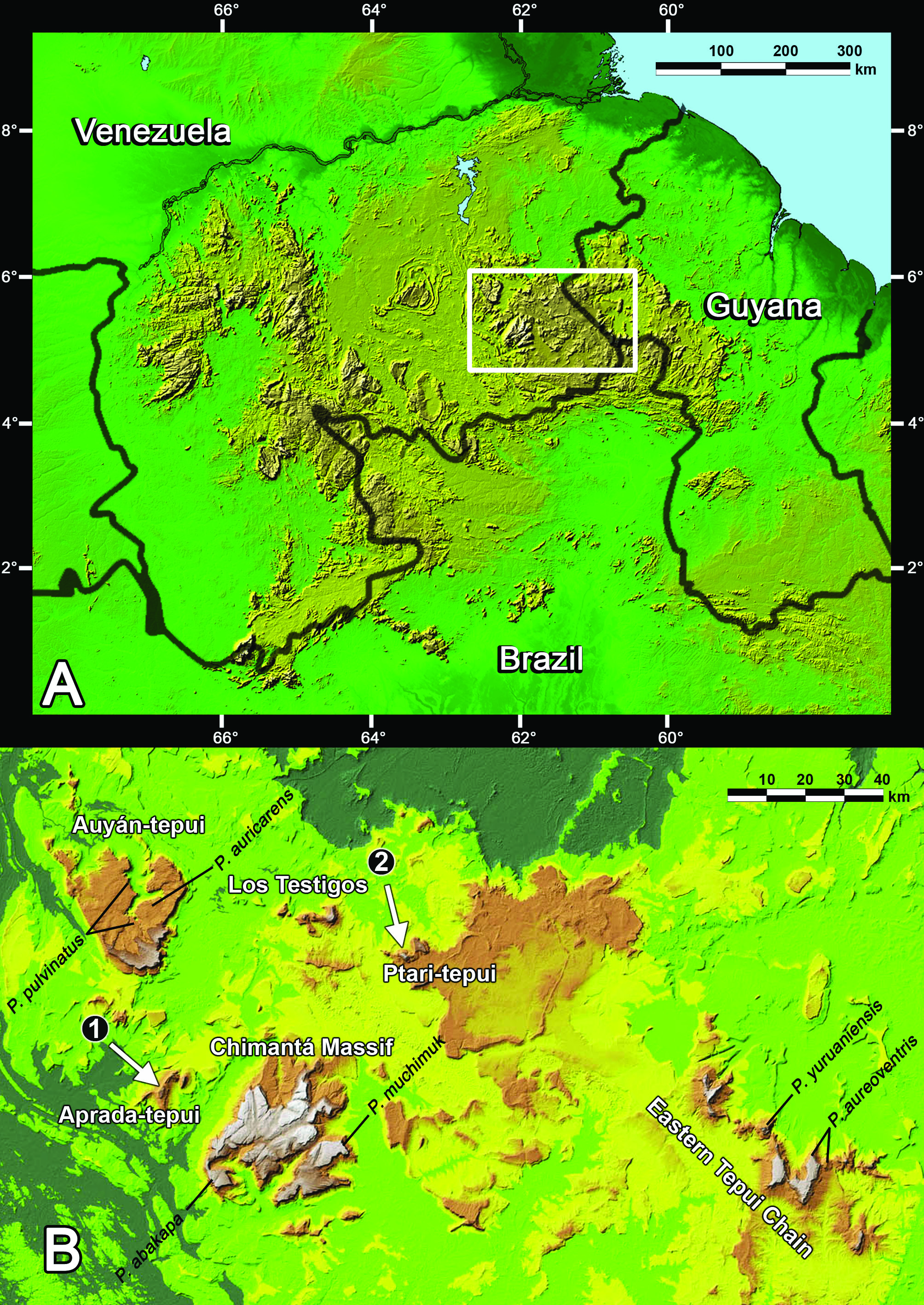
Pristimantis jamescameroni sp. nov. is only known from the summit of Aprada-tepui ( Figs 1B View Fig , 6A View Fig , 7 View Fig A-B), Bolívar state, Venezuela, where it occurs from 2557 to 2571 m elevation. Aprada-tepui lies in the Aprada massif ( Fig. 6A View Fig ), in the Chimantá subdistrict ( McDiarmid & Donnelly 2005). It is located ca. 22 km NW of the Chimantá massif and ca. 30 km S of Auyán-tepui (airline). According to the GPS, Aprada-tepui reaches a maximal elevation of about 2575 m above sea level. The summit area of Aprada-tepui is ca. 4.3 km 2, and is mainly covered by open rock vegetation and small islands of tepui forests ( Huber 1995, Fig. 7B View Fig ); it is characterized by a high number of small lakes ( Fig. 7B View Fig ) and some deep canyons.
Aside from the male holotype, which was collected by day on the ground while calling in a small shallow rock crevice, hidden by the vegetation, all specimens were collected under rocks. Other males were heard calling very sporadically from shallow rock crevices by day, but could not be located. As in most other tepui summit Pristimantis , the new species is not abundant and individuals are difficult to collect. The only other anuran reported from the summit of Aprada-tepui is Stefania satelles Señaris, Ayarzagüena & Gorzula, 1997 , which occurs in higher density and might partly feed on P. jamescameroni sp. nov. (pers. obs.).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pristimantinae |
|
Genus |
Pristimantis jamescameroni
| Kok, Philippe J. R. 2013 |
Pristimantis
| Hedges S. B. & Duellman W. E. & Heinicke M. P. 2008: 179 |
Eleutherodactylinae
| Gorzula & Señaris 1999: 55 |
Eleutherodactylus
| McDiarmid & Donnell, 2005: 514 |
Pristimantis
| Kok et al. 2012 |