
Allocapnia, RICKERI
|
publication ID |
https://doi.org/ 10.5281/zenodo.4760725 |
|
DOI |
https://doi.org/10.5281/zenodo.4763006 |
|
persistent identifier |
https://treatment.plazi.org/id/C86087E1-9326-F457-403C-FD3BFADFFC6E |
|
treatment provided by |
Felipe |
|
scientific name |
Allocapnia |
| status |
|
ALLOCAPNIA RICKERI View in CoL View at ENA GROUP
Ross & Ricker (1971) placed six species ( A. cunninghami Ross & Ricker 1971 ; A. perplexa Ross & Ricker 1971 ; A. rickeri Frison 1942 ; A. sandersoni Ricker 1952 ; A. stannardi Ross 1964 ; A. zola Ricker 1952 ) in this group and suggested, primarily on the basis of female characters, these form two “well- marked complexes of three species each”. The line of fusion between abdominal sterna 7-8 includes a sclerotized ridge in females of A. cunninghami , A. perplexa and A. zola , whereas females of A. rickeri , A. sandersoni and most A. stannardi lack such a structure. Subsequently, A. harperi Kirchner 1980 , was added to the second complex.
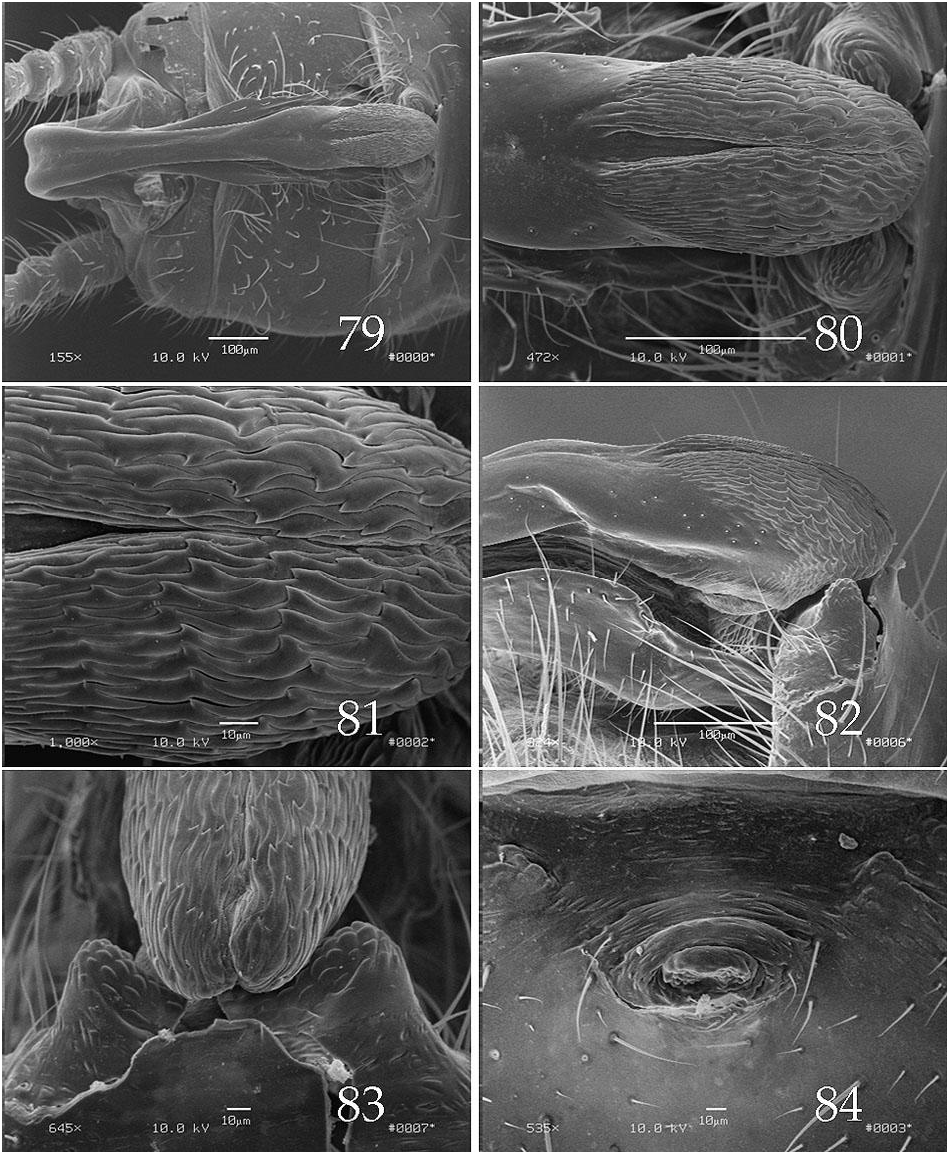
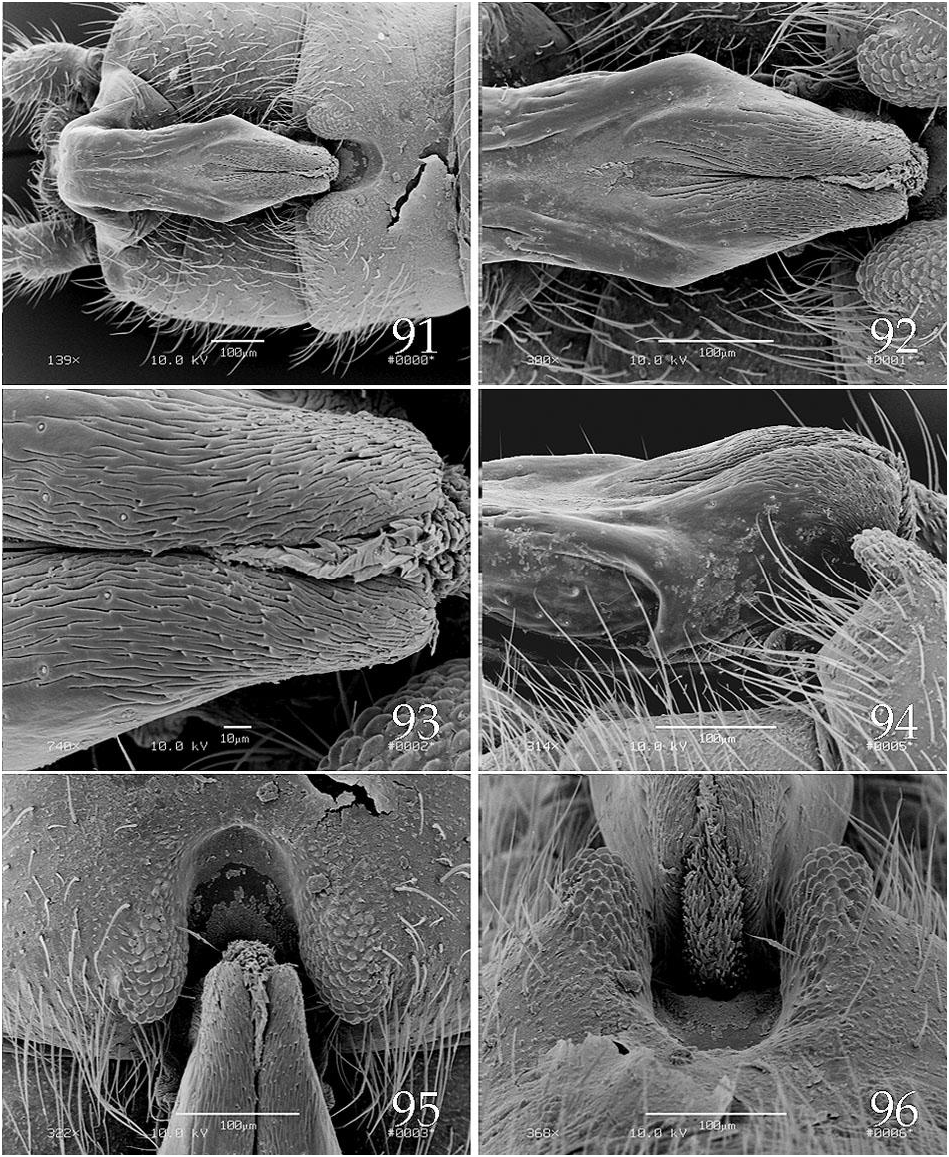
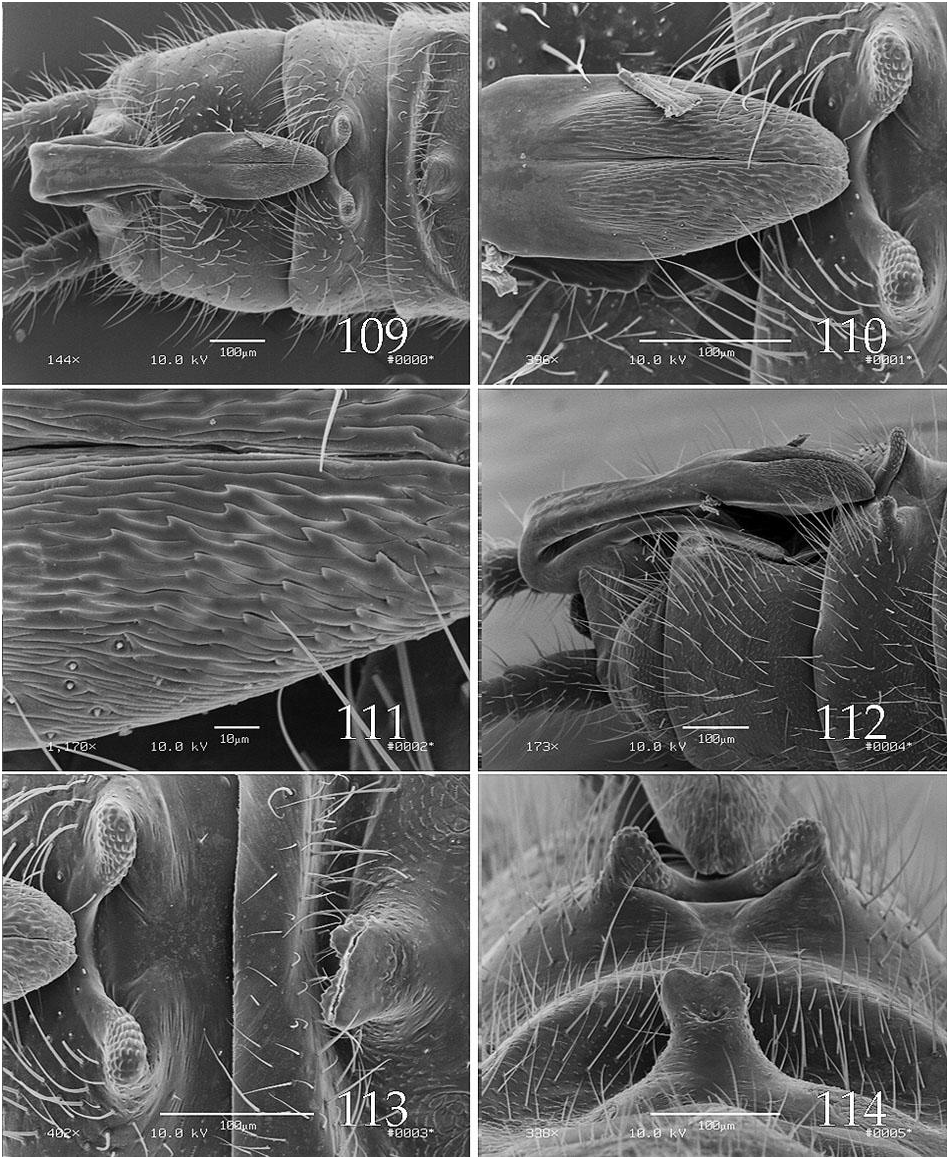
The epiproct upper limb for species in this group, is typified by a long basal segment and a relatively short, somewhat bulbous apical segment ( Figs. 79 View Figs , 85 View Figs , 91 View Figs , 97 View Figs ), a feature shared with members of the A. forbesi group. The apical segment bears a distal patch of densely packed, wave-like spikes ( Figs. 81 View Figs , 87 View Figs ) and the basal segment has a relatively broad, shallow longitudinal groove. The lower limb bears a distinct notch offsetting the apical spoon ( Fig. 100 View Figs ). Abdominal tergum 8, or 7 and 8, bear dorsal processes ( Figs. 90 View Figs , 95 View Figs , 112 View Figs ); those on tergum 8 are typically bilobed but in one species, A. zola , this structure is trilobed and a second process, also present on segment 8, is displaced toward the anterior segmental margin ( Fig. 115 View Figs ). In Ross & Ricker’s analysis, A. stannardi and A. rickeri were considered sister species with A. sandersoni more distantly related, and the other complex was presented as an unresolved trichotomy. The A. rickeri group shares “ancestor number 15” with A. jeanae Ross 1964 , in this analysis ( Ross & Ricker 1971).
We are unable to provide a definitive hypothesis for relationships within the A. rickeri group, but based primarily on tergal lobe variations, we could propose one subgroup in which the 8 th tergal lobe notch is at least 75 μm wide ( A. harperi , A. perplexa , A. rickeri , A. stannardi ; Figs. 90 View Figs , 96 View Figs , 101 View Figs , 114 View Figs ) and another in which the 8 th tergal lobe notch is no more than 45 μm wide ( A. cunninghami , A. sandersoni , A. zola ; Figs. 83 View Figs , 108 View Figs , 120 View Figs ). Within the first group, A. harperi , A. stannardi , and A. rickeri share ear-like lobes on the 8 th tergal process, whereas those of A. perplexa are not ear shaped, however we offer no resolution for the former trichotomy. In the A. cunninghami , A. zola , A. sandersoni subgroup, the former two species each share a small lobe on tergum 7 and also a small median projection on the 8 th tergal process which supports a sister group relationship between these two species. Unfortunately, we have no probable apomorphy which unites this latter subgroup.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.