
MACHAIRODONTINAE, Gill, 1872
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2005.00194.x |
|
persistent identifier |
https://treatment.plazi.org/id/CB5187EB-FFAE-FF90-4D08-FAA2BE47A7B5 |
|
treatment provided by |
Diego |
|
scientific name |
MACHAIRODONTINAE |
| status |
|
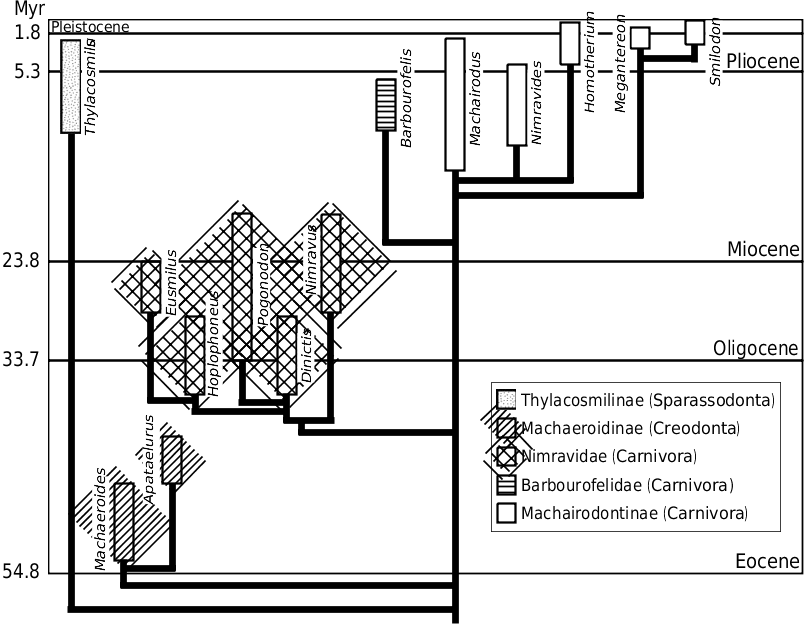
Machairodont felids, the ‘true’ sabrecats, are the best known of all sabretooths ( Matthew, 1901, 1910; Merriam & Stock, 1932; Kurtén, 1952; de Bonis, 1975; 1976; Martin, 1980; Rawn-Schatzinger, 1992; Turner & Antón, 1997). They first appeared during the late Miocene, are known from Africa, Eurasia, and the Americas, and went extinct at the end of the Pleistocene ( Fig. 1 View Figure 1 ) ( Martin, 1980; Turner & Antón, 1997). Two tribes are generally recognized, Homotherini and Smilodontini , representing the scimitar-toothed and dirk-toothed ecomorphs, respectively ( Kurtén, 1952; Martin, 1980). A third tribe is occasionally recognized, Metailurini , to include the ‘false sabretooths’ whose canines are intermediate between those of conical-toothed and sabretoothed felids ( Turner & Antón, 1997). Five species of scimitar-toothed [ Machairodus giganteus , Machairodus sp. (probably M. coloradensis ), Nimravides catocopis , Homotherium nestianus and Homotherium sp. (probably H. serum )] and three dirk-toothed [ Ischyrosmilus gracilis ( sensu Churcher, 1984; equivalent to Smilodon gracilis ), Smilodon fatalis and S. populator ] machairodonts were studied. Although it possesses sabre-like upper canines, Nimravides catocopis is often considered a very primitive homotherine because it lacks the derived skeletal features of sabretooths and possesses more robust lower canines (e.g. Turner & Antón, 1997; Martin, 1998b). Finally, the degree of variability in mandibular force profiles observed among dirktoothed machairodonts could represent a minimum because the smilodontines studied are closely related taxa. Future study of other smilodontine taxa should shed light on this possibility.
Mandibular force profiles
Although all machairodonts display a similar decrease in dorsoventral and labiolingual force values along the mandibular ramus, dirk-toothed and scimitar-toothed ecomorphs can be distinguished on the basis of their symphyseal properties, as was the case in nimravids (see above); however, the differences are much more subtle than in the Paleogene clade. Dorsoventrally ( Zx / L), the mandibular symphysis of homotherines is subequal or stronger than at P 4 M 1 ( Fig. 9 View Figure 9 ), while that of smilodontines is relatively weaker, being generally only slightly stronger than at P 3 P 4 ( Fig. 10). Labiolingually, the mandibular symphysis of homotherines is again subequal or stronger than at P 4 M 1, while that of smilodontines varies from slightly stronger to slightly weaker than at P 3 P 4 ( Figs 9 View Figure 9 , 10). This situation is the opposite of the pattern observed among nimravids, where dirk-toothed nimravids always have a symphyseal region stronger than the corpus at the carnassial while that of scimitar-toothed nimravids is much weaker, being only slightly stronger than at P 3 P 4.
The relative mandibular force profiles ( Zx / Zy) demonstrate more clearly the difference between homotherines and smilodontines. The mandibular ramus of smilodontines tends to becomes slightly rounder (decreasing Zx / Zy values) anteriorly, attaining a Zx / Zy minimum at P 3 P 4 ( Fig. 10), while that of homotherines remains relatively constant or tends to become gradually narrower (increasing Zx / Zy values) anteriorly, attaining a Zx / Zy maximum at P 3 P 4 ( Fig. 9 View Figure 9 ). Furthermore, the Zx / Zy canine values are higher than at P 3 P 4 in smilodontines but the opposite is observed in homotherines. The increase is so great among smilodontines that their mandibular symphysis is much better buttressed dorsoventrally, or deeper, than that of homotherines; in other words, the Zx / Zy canine values of dirktoothed machairodonts are higher than those of scimitar-toothed machairodonts ( Figs 9 View Figure 9 , 10). Surprisingly, the best-known sabretooth, Smilodon fatalis from the
Dorsoventral force Labiolingual force Relative force (Log Zx/L) (Log Zy/L) ( Zx/Zy)
2.29 2.19
Machairodus giganteus ( L = 23.94 cm)
Machairodus sp. ( L = 20.77 cm)
Nimravides catocopis ( L = 20.73 cm)
Dorsoventral force Labiolingual force Relative force ( Log Zx/L) ( Log Zy/L) ( Zx/Zy)
Rancho La Brea deposits, appears to differ from the typical dirk-toothed machairodont profile in having a symphyseal region only slightly better dorsoventrally buttressed than at P 3 P 4 ( Zx / Zy canine = 1.89 in S. fatalis contra Zx / Zy canine > 2.48 in other smilodontines).
Interpretation
Assuming similarity of cortical bone thickness and of safety factors, comparison of the Zx / L profiles of machairodonts with those of modern felids indicates machairodonts had more powerful bites than extant felids of similar mandibular length ( Table 1). Indeed, comparison at the P 4 M 1 interdental gap reveals that Homotherium had a bite force 38–62% greater than Panthera leo , while the studied smilodontines and Nimravides had a bite force from 2% to 51% greater than P. leo . Machairodus is the only machairodont for which the estimated bite force is lower than that of a similar-sized felid, being only 78–84% the bite of the lion. Of course, these values most assuredly underestimate the actual bite forces of these creatures because, as Akersten (1985) showed, the mandibular cortical bone thickness of S. fatalis was much greater than in P. leo .
The fact that the dorsoventral and labiolingual force profiles of dirk-toothed and scimitar-toothed machairodonts are the opposite of those observed in nimravid ecomorphs could be related to the labial inclination of the mandibular corpus near the cheek teeth observed in smilodontines, a condition similar to that of barbourofelids. In contrast, the orientation of the mandibular corpus in homotherines remains constant and the cheek teeth are in the median plane of the ramus (pers. observ.). Owing to the labial rotation of the ramus, the labiolingual diameter of the corpus appears greater in smilodontines than in homotherines, resulting in higher force values along the ramus relative to the mandibular symphysis. These inflated values could explain why force values at the canine in smilodontines are only slightly higher than at P 3 P 4, as seen in barbourofelids, while those in homotherines are relatively much higher. These differences are recognizable in the Zx / Zy profiles, where smilodontines appear to have slightly rounder mandibles (lower Zx / Zy values) than homotherines ( Figs 9 View Figure 9 , 10). Therefore, it is highly probable that the feeding habits of machairodont dirk-toothed and scimitartoothed ecomorphs did not differ drastically from those of their respective nonfelid sabretooth ecomorphs despite their differing mandibular force profiles.
The Zx / Zy canine values further support the similarity of feeding behaviour between the respective machairodont and nonmachairodont ecomorphs. Although all machairodonts possess high Zx / Zy canine values relative to extant carnivorans, the symphysis of smilodontines tends to be better dorsoventrally buttressed (higher Zx / Zy canine values) than that of homotherines (difference not statistically significant due to the low Zx / Zy canine values of S. fatalis ). Such results suggest that dirk-toothed smilodontines used their sabre bite on better-restrained prey than scimitar-toothed homotherines, thus reducing the amount of labiolingual and torsional stresses exerted at the symphysis. This is not surprising given the documented postcranial differences between the two taxa ( Kurtén, 1952; Martin, 1980; Anyonge, 1996). Smilodontines would have been able to immobilize prey with their powerful forelimbs prior to delivering a powerful sabre bite while homotherines could have delivered shallow, slashing bites during pursuit to weaken their prey, immobilize it with their robust incisor battery (Biknevicius et al., 1996), and used their sabres as a ‘coup de grâce’ to finish their prey after it had fallen (see Antón & Galobart, 1999).
The lower Zx / Zy canine value of S. fatalis relative to other smilodontines (and sabretooths in general) is perplexing, particularly given that this value is very close to that of the extant Neofelis nebulosa . Because S. fatalis is a full-fledged dirk-toothed ecomorph, there is no obvious morphological reason why this taxon should have been less efficient at immobilizing prey than other sabretooths, yet its low Zx / Zy canine value indicates that greater torsional stresses were exerted at the symphysis. The fact that all studied specimens of S. fatalis are from the Rancho La Brea tar pits, a unique predator trap setting, could be related to the peculiarity of these results. The high frequency of tooth fracture in Rancho La Brea predators, including S. fatalis , was interpreted as evidence for increased carcass utilization in response to intense predator competition near the tar pits ( Van Valkenburgh & Hertel, 1993). More rapid and less careful feeding would generate greater torsional stresses at the symphysis, especially because incisors are the most commonly used teeth to acquire flesh from a carcass ( Van Valkenburgh, 1996). These greater torsional stresses would, thus, require S. fatalis to develop a less dorsoventrally buttressed symphysis than sabretooths hunting under normal (lower) competitive conditions, which could feed more slowly and carefully. One way to test the validity of this hypothesis is to compare the mandibular force profiles of S. fatalis individuals collected from Rancho La Brea with individuals of the same species found elsewhere in North America to see if their Zx / Zy canine values differ. If the mandibular force profiles of North American S. fatalis individuals prove to be closer to those of other smilodontines rather than to individuals from Rancho La Brea, then support for the influence of the peculiar Californian settings on the feeding behaviour of the sabretoothed predator will be gained. In contrast, if all S. fatalis individuals possess lower Zx / Zy canine values than other smilodontines, then different causes need to be invoked to explain the different feeding behaviour of this taxon.
Finally, the incipient-sabretooth Nimravides catocopis appears to have hunted in a true sabretooth fashion. Unlike the nimravid N. brachyops , which possessed force profiles intermediate between those of true sabretooths and conical-toothed felids but similar to those of Neofelis nebulosa , the mandibular force profiles of Nimravides are undistinguishable from those of more derived homotherines. Consequently, in spite of its poorly developed sabretooth craniodental adaptations, Nimravides probably behaved as a true, yet primitive sabretooth.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.