
Oligosarcus itau, Mirande, Juan Marcos, Aguilera, Gastón & Azpelicueta, María De Las Mercedes, 2011
|
publication ID |
https://doi.org/10.5281/zenodo.201381 |
|
DOI |
https://doi.org/10.5281/zenodo.6191724 |
|
persistent identifier |
https://treatment.plazi.org/id/CF555B6D-FF97-B845-D7E1-62B1FC90F4E9 |
|
treatment provided by |
Plazi |
|
scientific name |
Oligosarcus itau |
| status |
sp. nov. |
Oligosarcus itau View in CoL , new species
( Figs. 1–2 View FIGURE 1 View FIGURE 2 , Table 1)
“ Astyanacinus ” sp., Mirande, 2008
Characidae View in CoL n. gen., n. sp., Mirande, 2009 Oligosarcus View in CoL sp., Mirande, 2010
Holotype. CI-FML 3856, male, 62.1 mm SL, Argentina, Salta, General San Martín, Río Bermejo basin, small tributary to Río Itau , near Campo Largo, 22º 01' 34'' S, 63º 56' 06'' W, 690 m a.s.l. coll. J. M. Mirande, G. Aguilera & C. Aguirre. November 2005.
Paratypes. CI-FML 3857, 1 ex., female, 66.2 mm SL, collected with holotype. CI-FML 3858, 1 ex., male, C&S, 61.4 mm SL, collected with holotype.
Diagnosis. Distinguished from remaining species of Oligosarcus by presence of two well distinct rows of premaxillary teeth (vs. one row composed by both large and small teeth) ( Fig. 3 View FIGURE 3 ). One C&S specimen of Oligosarcus itau has four unbranched dorsal-fin rays, a feature not found in other characids; however, presence or absence of very small anteriormost dorsal-fin ray could not be evaluated in alcohol-preserved specimens. The new species further distinguishes from all remaining species of genus excepting O. pintoi by presence of pentacuspidate dentary teeth (vs. conical to tricuspidate). Oligosarcus itau further distinguishes from O. pintoi by presence of 8–13 maxillary teeth (vs. 15–23) ( Fig. 3 View FIGURE 3 ), 21–23 branched anal-fin rays (vs. 24–28), 18–19 circumpeduncular scales (vs. 13– 16), 41–42 lateral-line scales (vs. 36–40), presence of two pairs of uroneurals (vs. one), presence of a well-developed temporal fossa ( Fig. 4 View FIGURE 4 )(vs. absent; Fig. 5 View FIGURE 5 ), and possession of a distally expanded sphenotic spine ( Fig. 4 View FIGURE 4 )(vs. distally notched; Fig. 5 View FIGURE 5 ).
Further distinguished from Oligosarcus bolivianus , also inhabiting Río Bermejo basin, by shorter maxilla, not reaching middle length of eye (vs. reaching, or almost reaching vertical through posterior margin of eye), lower number of ectopterygoid teeth (3–4 vs. 9–11), lower number of lateral-line scales (41–43 vs. 49–55), transverse (9/ 5–7 vs. 11/8), and circumpeduncular scales (18–19 vs. 21–23), and fewer number of maxillary teeth (8–13 vs. 15– 24). Oligosarcus itau distinguishes from O. platensis n. comb. (see Discussion) by fewer anal-fin rays (21–23 vs. 27) and maxillary teeth (8–13 vs. 16) and by dentary teeth gradually decreasing in size (vs. two anterior dentary teeth conspicuously longer than posterior ones).
Description. Morphometrics of holotype and two paratypes are presented in Table 1. Body moderately deep (36.3–37.5 % SL), with maximum depth at dorsal-fin origin. Dorsal profile of snout slightly convex and straight to tip of supraoccipital process; slightly convex to dorsal-fin origin; almost straight from this point to caudal peduncle; gently concave along caudal peduncle to base of caudal-fin rays. Ventral profile of body gently convex from lower-jaw tip to pelvic-fin origin; straight to anal-fin origin; straight along base of anal fin, and gently concave under caudal peduncle. Ventral portion of body between pectoral and pelvic fin origins transversely rounded; ventral portion of body between origins of pelvic and anal fins more compressed laterally.
Dorsal-fin origin slightly posterior to middle standard length (predorsal distance 54.7–57.0 % SL). Pelvic-fin origin situated anterior to vertical through dorsal-fin origin. Anal-fin origin situated at vertical through base of posteriormost dorsal-fin rays. Tip of pectoral fin reaching pelvic-fin origin when adpressed against body; tip of pelvic fin not reaching anal-fin origin.
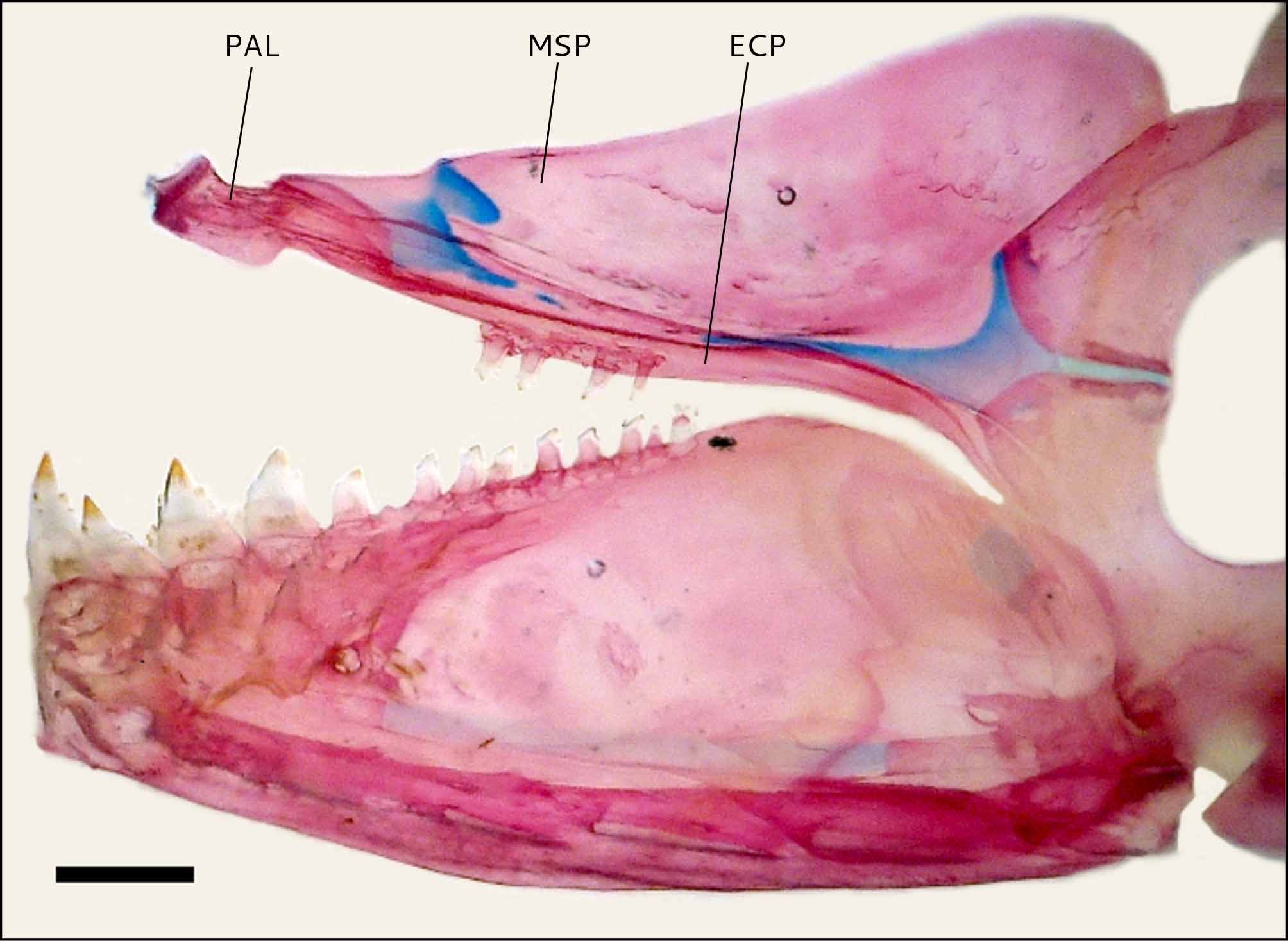
Mouth terminal, situated at level of inferior third of eye; maxilla following a rather continuous line with premaxilla, extended posteriorly to anterior third of orbit. Premaxilla bearing two series of teeth. Outer row with 4(3*) tricuspidate teeth, with very small lateral cusps. Inner row with 5 narrow teeth with a central cusp well developed, and lateral cusps smaller; symphysial tooth very slender and long, with 4 cusps; second and third teeth slightly wider with 5 cusps; fourth and fifth teeth with 3–4 cusps; medial three teeth on inner premaxillary row separated from each other by visible spaces. Laminar process of maxilla rather long, with 8–13 conical or tricuspidate teeth, reaching 3/4 of its length ( Fig. 3 View FIGURE 3 ); ligamentum primordiale attached on horizontal line through first maxillary tooth base. Dentary with maximum depth approximately at posterior fourth of its length, bearing 4 pentacuspidate teeth followed by 10 smaller tricuspidate or conical teeth ( Fig. 6 View FIGURE 6 ). Lateral cusps of dentary teeth much smaller than medial one. Ectopterygoid with 3–4 teeth ( Fig. 6 View FIGURE 6 ).
Eye moderately large, longer than snout. Third infraorbital not contacting laterosensory canal of preopercle.
Dorsal fin with iv,9 rays; first ray visible only in C&S specimen; distal margin of dorsal fin straight, with last unbranched and first branched dorsal-fin rays longest. Anal fin with v-vi,21(1*), 23(2) rays. Caudal fin with principal rays i,17,i. Pectoral fin with i,12, i,13(*), or i,14 rays. Pelvic fin with i,7 rays.
Scales cycloid, without circuli on posterior field. Lateral line complete with 41(1*) or 42(2) perforated scales. Scales between dorsal-fin origin and lateral line 9(3*); scales between lateral line and pelvic-fin origin 5(1*), 6(1), or 7(1). Scales around caudal peduncle 18(2*) or 19(1). Predorsal scales 16–17. One row of 10–14 scales forming a sheath covering base of anterior anal-fin rays. Few scales covering only proximal region of caudal-fin lobes.
First branchial arch with 22–23 gill rakers: 8–9 on epibranchial, 1 on cartilage, 10–11 on ceratobranchial, and 2 on hypobranchial; posterior edge of first epibranchial with a second row of 3–5 gill rakers. Gill rakers on first ceratobranchial with broad denticles especially on anterior and posterior edges. Thirty-six vertebrae (18 precaudal and 18 caudal). Five supraneurals, 12 pairs of ribs. Caudal fin with 12 dorsal and 10 ventral procurrent rays.
Sexual dimorphism. S mall anal-fin lobe formed by last unbranched and first 1–2 branched rays in both sexes, pointed in female and rounded in males. Males with hooks on last unbranched anal-fin ray and posterior branch of first 9–11 branched anal-fin rays; one pair of hooks per segment. Males bearing hooks on pelvic-fin rays 2–5, one pair of hooks per segment. One small hook at tip of caudal fin of one C&S male specimen. No gill-derived gland observed in males or females.
Colour in life. Body silvery, darker dorsally. Lateral band faint. Two vertically elongated humeral spots, anterior one well defined and narrow and posterior one faint. Caudal spot extended to end of median caudal-fin rays. Dorsum of head and snout dark. Cheek and opercle silvery and finely dotted with melanophores; grey maxilla. Dorsal region of eye red or brown. Fins yellow proximally and red distally ( Fig. 1 View FIGURE 1 ).
Colour in alcohol. Body yellowish, darker dorsally. Dorsum of head dark with evident chromatophores scattered on cheek and opercle. Scales from upper half of flank more densely pigmented, especially in their distal margin, giving a reticulated aspect. First humeral spot vertically elongated, anteriorly situated on flank, with its ventral end almost contacting posterior margin of opercular flap. A rather thin lateral band on flank extended from anterior humeral spot, through a diffuse posterior humeral spot and caudal peduncle to end of middle caudal-fin rays. A dark, rhomboidal spot present at caudal peduncle ( Fig. 2 View FIGURE 2 ).
Distribution. Oligosarcus itau is only known from an unnamed stream in the road to Campo Largo, General San Martín, Salta, Argentina; this stream is an affluent of the Río Itau , Río Bermejo basin ( Fig. 7 View FIGURE 7 ).
Conservation notes. Specimens were found in a rainforest stream with riffles and pools of one or two meters with bottom ranging from gravel to large rocks ( Fig. 8 View FIGURE 8 ). The stomach of one C&S specimen contained mainly allochthonous insects. The stream in which this species was collected flows through the piedmont rainforest of north-western Argentina; these environments are highly endangered and are disappearing due to human activities. The particular area where this species inhabits is concessioned to a hydrocarbon exploitation company and it is subjected to permanent modifications of their water courses; this and the apparently natural scarcity of specimens of this species in its only known locality make it highly vulnerable. Despite the relatively high collection efforts in the basin, this species was found only in this stream, and probably it constitutes a microendemism.
Etymology. The specific name itau refers to the Río Itau . This word probably derives from the guaraní words “ itá ”, meaning stone, and “y” (pronounced as the German ü in Guaraní language), meaning water. A noun in apposition.
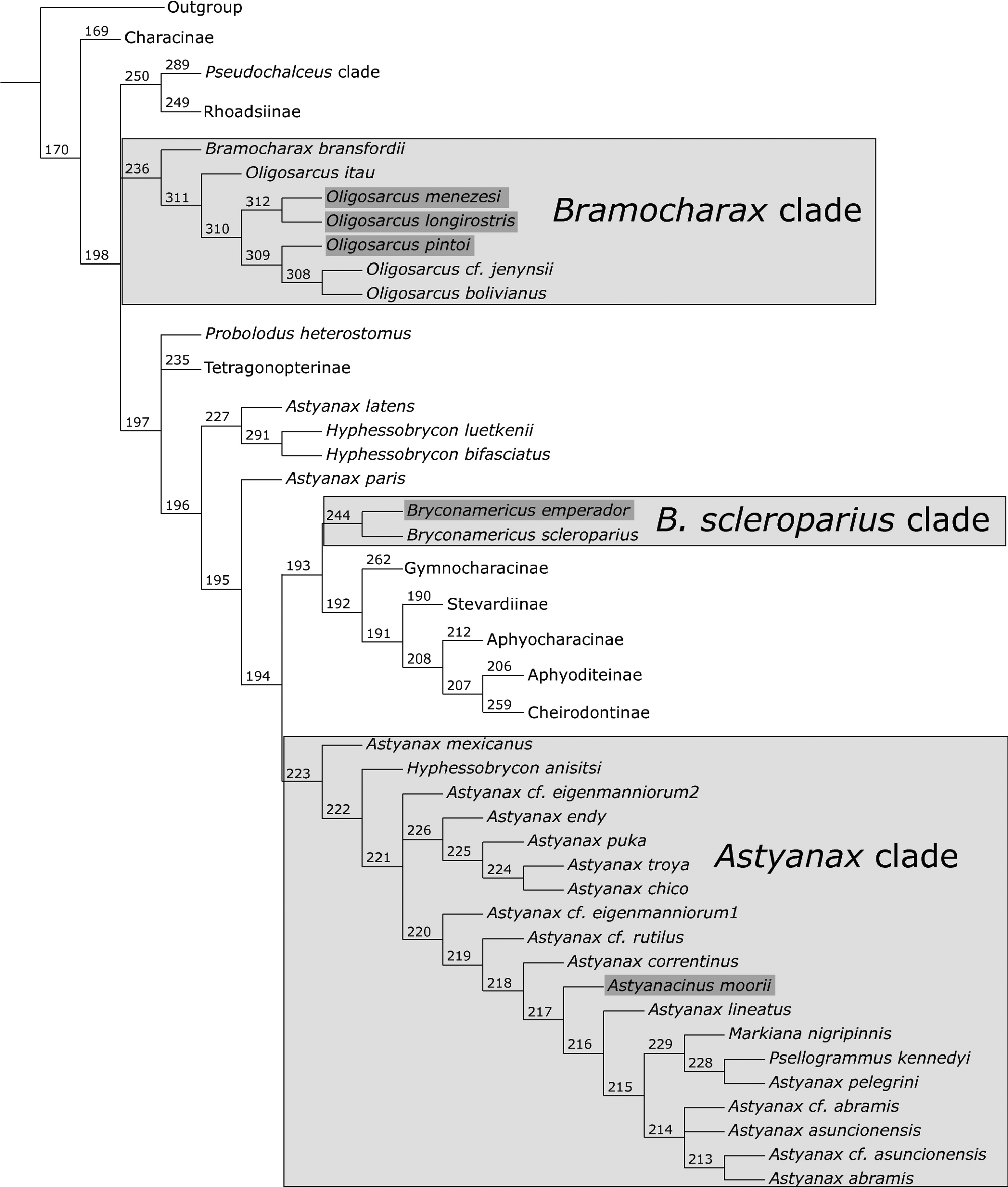
Phylogenetic results. The phylogenetic hypothesis proposed herein is the strict consensus of the most parsimonious trees obtained between the 8th and 14th values of k (9.00–16.44) in the analyses under implied weighting. This phylogeny is much similar to that proposed by Mirande (2010), and the discussion is principally focused on the relationships of Oligosarcus and Astyanacinus ( Fig. 9 View FIGURE 9 ). Characters and states listed correspond to that described by Mirande (2010), excepting for character 366 (see below).
The resolution of the internal relationships of Oligosarcus was not a primary objective of this paper; however, it is noticeable that O. itau n. sp. is the sister taxon of the remaining analyzed species of the genus in all the analyses performed. The monophyly of Oligosarcus is well supported and stable across all the analyses, both under implied weighting and self-weighted optimization ( Fig. 9 View FIGURE 9 ).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |