
Parkermavella columnaris, Martino & Taylor & Gordon & Liow, 2017
|
publication ID |
https://doi.org/10.5852/ejt.2017.345 |
|
publication LSID |
lsid:zoobank.org:pub:3F002D75-2A32-4300-BCE1-683C7311B70C |
|
DOI |
https://doi.org/10.5281/zenodo.3850221 |
|
persistent identifier |
https://treatment.plazi.org/id/D3D59208-F73C-4A9A-960A-12A042FCB8EE |
|
taxon LSID |
lsid:zoobank.org:act:D3D59208-F73C-4A9A-960A-12A042FCB8EE |
|
treatment provided by |
Carolina |
|
scientific name |
Parkermavella columnaris |
| status |
sp. nov. |
Parkermavella columnaris sp. nov.
urn:lsid:zoobank.org:act:
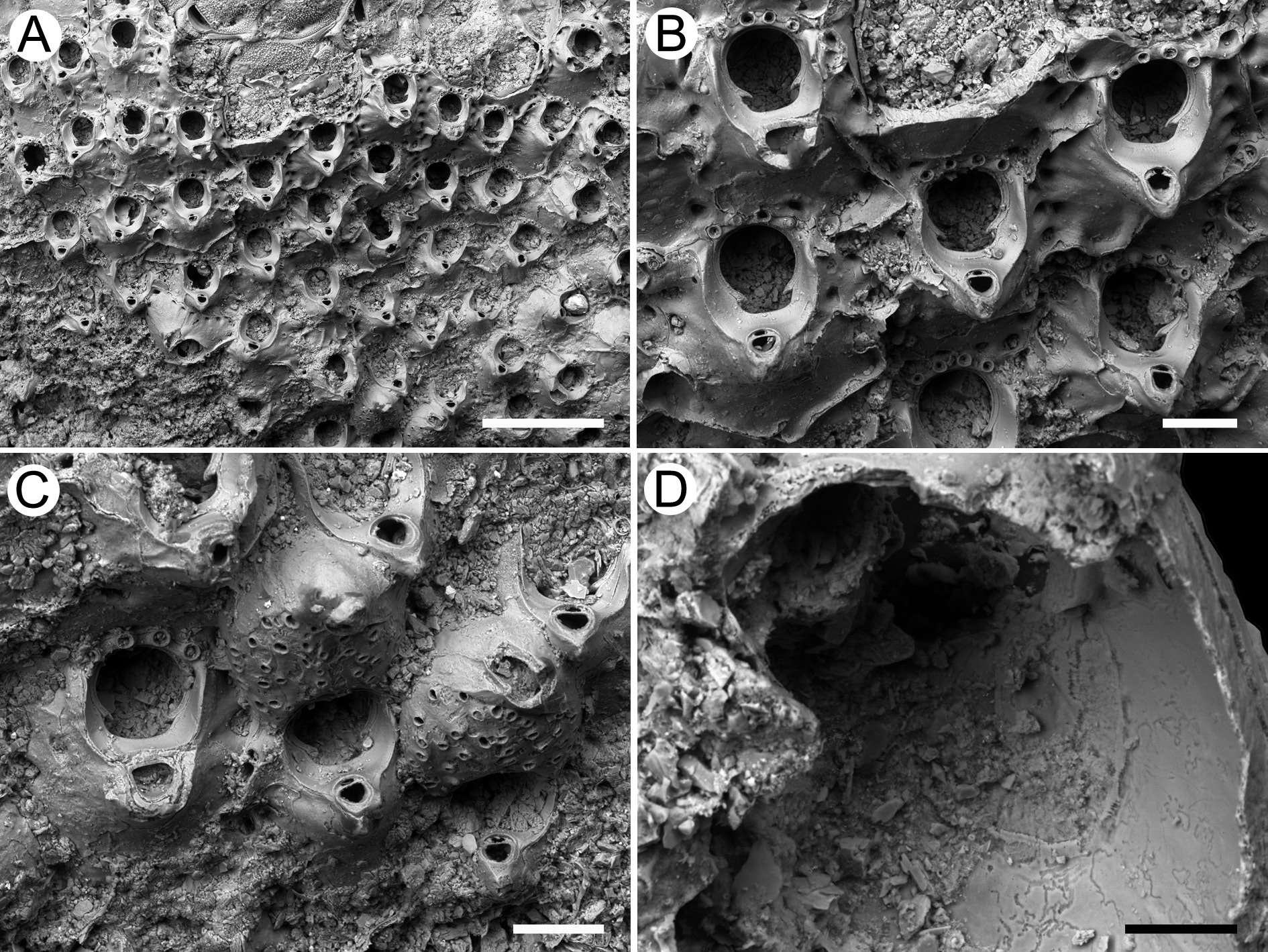
Figs 2–3 View Fig View Fig , Tables 2–3 View Table 2
Diagnosis
Colony encrusting. Autozooids polygonal. Frontal shield partially umbonuloid, smooth and imperforate centrally, with a continuous row of marginal areolar pores. Orifice broadly cleithridiate (keyhole shaped), with condyles delimiting the arched anter from a broad sinus. Distal oral spines present. A columnar structure bearing an adventitious suboral avicularium with a complete crossbar constantly present. Ooecium lepralielliform; ectooecium calcified, smooth, with pseudopores; secondary calcification of the frontal shield of the distal zooid covering the distal half of the ooecium and bearing an avicularium similar to the suboral one.
Etymology
From the Latin columnaris , -e, pillar-like, referring to the columnar peristome bearing the suboral avicularium.
Material examined
Holotype
NEW ZEALAND: Recent ,? Cook Strait ( NIWA 97418 View Materials ).
Paratypes
NEW ZEALAND: Upper Kai-Iwi Shellbed, Castlecliff Beach, Castlecliffian, Pleistocene ( NHMUK PI BZ 7832, NHMUK PI BZ 7833).
Description
Colony encrusting, small in fossil colonies, up to 33 mm in diameter in Recent holotype colony.Ancestrula and early astogeny not observed. Autozooids arranged in parallel rows ( Figs 2A View Fig , 3A View Fig ), varying in shape, commonly rectangular, sometimes hexagonal or irregularly polygonal, longer than broad (mean L/W = 1.29). Frontal shield partially umbonuloid, convex centrally, flat proximally, smooth and imperforate apart from a continuous row of numerous large (20–35 μm), evenly distributed areolar pores, usually 7−9 along lateral and distal margins, oval to circular, funnel-shaped or sloping outward. Sometimes, one or two additional areolar pores form a second row in distolateral corners of zooids. Orifice broadly cleithridiate ( Figs 2B View Fig , 3B View Fig ). Two small, pointed, downward-directed condyles placed at two thirds of orifice length, separating an arched anter from a broad, shallow, bowl-shaped sinus. Two, three or four oral spine bases (10–22 μm in diameter) placed on distal orificial margin, hidden in ovicellate zooids. Peristome developed as two lateral wings, rising proximally, forming a somewhat columnar structure, rising 150–160 μm from surface of frontal shield in lateral view, always bearing a suboral avicularium ( Fig. 2C View Fig ). Peristome hiding primary orifice in frontal view. Adventitious avicularium suboral, small, oval, upward-directed, sloping distally, rostrum rounded and with a complete crossbar ( Figs 2B View Fig , 3 View Fig B–C). Ooecia of lepralielliform type, prominent and globular ( Figs 2A View Fig , 3C View Fig ). Ectooecium calcified, smooth, centrally perforated, pseudopores bordered by a raised margin, varying in size (10–30 μm) and form from circular to bean-shaped. Secondary calcification from frontal shield of distal zooid covers distal half of ooecium, forming a raised crest bearing an avicularium similar in size and shape to suboral avicularium. The two avicularia aligned, but oppositely directed.
Remarks
Parkermavella was introduced by Gordon & d’Hondt (1997) for Schizomavella -like species with frontal shields imperforate apart from marginal areolar pores. In the diagnosis of the genus ( Gordon & d’Hondt 1997: 17), the zooids are defined as lepralioid; however, new observations have revealed, for at least some of the species, the presence of an umbonuloid area of variable size. For instance, in P. curvata (Uttley & Bullivant, 1972) the umbonuloid area is large, occupying most of the frontal shield, while in P. punctigera (MacGillivray, 1883) and P. virago ( Gordon, 1989) it is more reduced, slightly smaller than the orifice in the former species and about the size of the orifice in the latter species (D.P.G., personal observation). In P. columnaris sp. nov. the umbonuloid area is small ( Figs 2D View Fig , 3D View Fig ), 120 μm long by 95 μm wide, and possibly coincides with the overlying avicularian cystid.
This genus is moderately diverse in New Zealand waters at the present day, with seven species formally described from the region and several others awaiting description (D.P.G., unpublished data). The new species is easily distinguishable from all other known species of Parkermavella by its unique suite of morphological characters, specifically the smooth frontal shield (nodular-tubercular in many other species), the broadly keyhole-shaped orifice with a large and concave sinus (narrow and/or straight in the others), and the unique columnar structure bearing the suboral avicularium. P. virago ( Gordon, 1989) has a similar smooth frontal shield and development of the secondary calcification of the distal zooid covering the ooecia, but differs in the more numerous, smaller avicularia suborally and in the secondary calcification.
The sole Recent colony of P. columnaris sp. nov. differs from the fossil ones in having slightly larger zooids, orifices and ooecia, although the L/W ratio is identical ( Tables 2–3 View Table 2 ).
Distribution
The oldest geological record of this species dates back to the Nukumaruan (2.29–2.08 Ma, Pleistocene) of the Wanganui Basin. A small number of colonies were found encrusting bivalve shells from the Nukumaru Limestone, and on bivalve shells of the Castlecliffian Upper Kai-Iwi Shellbed (0.68–0.62 Ma, Pleistocene). The Recent colony of Parkermavella columnaris sp. nov. housed in the NIWA collection (i.e., the holotype) unfortunately lacks collection data. It encrusts a 75 mm-long valve of the pholadid bivalve Barnea similis (Gray, 1835) , along with the bryozoans Antarctothoa bathamae (Ryland & Gordon, 1977) , A. tongima (Ryland & Gordon, 1977) , Caberea rostrata Busk, 1884 , Chaperiopsis lanceola Hayward & Thorpe, 1988 , Cornuticella taurina (Busk, 1852) , Opaeophora lepida (Hincks, 1881) , Orthoscuticella fissurata (Levinsen, 1909) and Tricellaria aculeata (d’Orbigny, 1842) . The known distributions of these species ( Gordon 1986, 1989) overlap or coincide only in greater Cook Strait, a provenance correlated with the presence there of rocks bored by pholadids; hence we conclude that the holotype specimen came from a locality in the greater Cook Strait area.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Smittinoidea |
|
Family |
|
|
Genus |