
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/10.1671/0272-4634(2000)020[0001:adreft]2.0.co;2 |
|
DOI |
https://doi.org/10.5281/zenodo.3812839 |
|
persistent identifier |
https://treatment.plazi.org/id/D32F092B-552A-904C-4689-FD6C127AF072 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
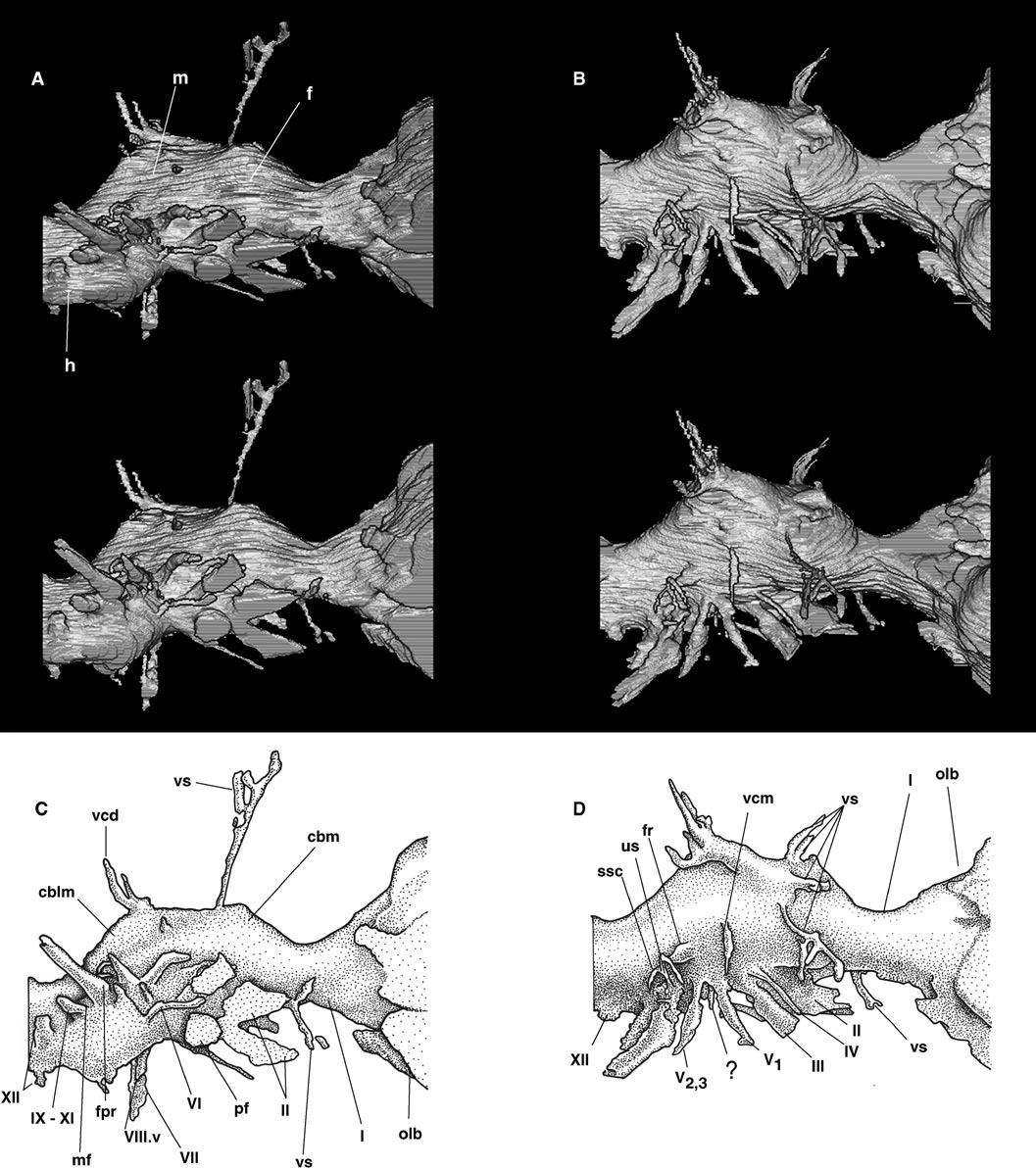
The skull of a mature specimen of Tyrannosaurus rex
in the Field Museum of Natural History ( FMNH PR2081, ‘‘Sue’’ ) was recently subjected to highresolution computed tomographic (CT) analysis, and an endocast was generated without physical damage to the skull. It improves upon endocasts made through hemisection for the tyrannosaurids Tyrannosaurus ( Osborn, 1912) and Tarbosaurus ( Maleev, 1965) by adding details of the inner ear and nerve and vascular pathways not visible through these earlier destructive methods. In particular, it reveals greatly enlarged olfactory bulbs, which suggests high olfactory acuity.
Methods —The CT scanner generated 748 coronal ‘‘slices’’ through the skull, from the tip of the snout to the back of the skull. These 2 mm thick slices were generated with a Minatron 250 scanner built by Scientific Measurement Systems, Inc., and operated at the Santa Susana Field Laboratory (SSFL) of Rocketdyne Division, Boeing, Inc., Chatsworth, California. The skull was wrapped in aluminum foil and encased in two different grades of injectable polyurethane foam made by InstaFoam Products, Inc., Division of Flexible Products Co.: a onepound foam to coat very fragile regions of the skull and a fourpound foam for the remainder. A wooden crate was built around the partiallyfoamed skull and fourpound foam was used to fill all voids. This crate was rotated 90 degrees at SSFL such that the occipital plate was parallel to the ground and the nose pointed up.
An ideal method for producing a digital endocast would be to ‘‘remove’’ the bone from the data set, leaving an image of the matrix, including the matrix filling the endocranial cavity and associated vascular and nervous structures. While most matrix filling the skull was a sandy siltstone, its density varied. Some portions of the endocranial cavity and nerve canals were completely empty, and others filled with matrix nearly as dense as the bone itself. Moreover, most of the matrix portion of the data set was noisy and overlapped the bone portion. For this reason, digital ‘‘removal’’ of bone was not possible without also removing much of the matrix. This confounded attempts to synthetically generate smooth, continuous surfaces. Features of the central nervous system and otic capsule were traced by hand on horizontal slices through the braincase in Adobe Photoshop 4.0 (Adobe Systems, Inc., San Jose, California), using horizontallyoriented slices through the braincase. Tracings were assembled into a threedimensional image using VoxBlast 2.2 (VayTek, Inc, Fairfield, Iowa).
The skull was scanned before preparation began on the braincase. This increased the stability of the specimen during transport and scanning. However, as preparation of the braincase began long after the digital endocast was generated (indeed, portions of the braincase remain covered with matrix), I was unable to crosscheck cranial nerve tracts on the external braincase wall for several months, with the exception of the tenth through twelfth, which exit ventrolateral to the occipital condyle. External foramina can now be seen for the seventh and first five cranial nerves on the right side.
Gross Morphology —Nonavian reptilian endocasts are only coarse approximations of the shape of the brain ( Edinger, 1929; Hopson, 1979). The endocast is not perfectly symmetrical, as the nerve and vascular passages on the left were filled with denser matrix than on the right and were not as easily traced. But divisions between the fore, mid, and hindbrain can be seen on the endocast ( Fig. 1A View FIGURE 1 ). The cerebellum would have been within an expanded region behind the cerebrum. The medulla is represented by the narrow tubular portion of the endocast behind the cerebellum. The position of the otic capsule is betrayed by a lateral constriction in the cerebellar region, with the thin floccular process projecting posteroventrolaterally and visible between the delicate semicircular canals.
The endocranial cavity is 28.2 cm long from the foramen magnum to the anterior margin of the orbitosphenoids. This is 8 cm longer than the endocast described by Osborn (1912), but Osborn was using the braincase of a small and, presumably, immature T. rex ( American Museum of Natural History 5029 View Materials ). The olfactory tract, with bulbs, extends anteriorly for approximately 20 cm. The cerebral portion of the endocast is approximately 9 cm wide and 7 cm deep.
Endocranial volume in nonmaniraptoran theropods—including tyrannosaurids—falls within the predicted range for nonavian reptiles of appropriate body mass ( Jerison, 1973). More basal theropods and nontheropod dinosaurs retained the plesiomorphic arrangement of a large cerebellum projecting dorsally over the roof of the forebrain ( Marsh, 1896; Gilmore, 1920; Madsen, 1976; Rogers, 1998, 1999). The forebrain of T. rex is large relative to its counterpart in noncoelurosaurian theropods, but is relatively small compared with those of closer relatives of birds, such as Troodon ( Russell, 1969) .
All twelve cranial nerves are visible, though not always on both sides ( Fig. 1 View FIGURE 1 ). The olfactory tracts ( I) project through a common opening between the orbitosphenoids anteriorly, and are visible on the endocast as a thick process at the front of the endocast. The optic nerves ( II) both diverge from a single large midline opening in the laterosphenoids in tyrannosaurids ( Osborn, 1912; Russell, 1970), and the mold of this opening is visible on the digital endocast. A thin bony wall separates the optic foramen and pituitary fossa. The trochlear nerve ( IV) exits the forebrain dorsal to the oculomotor ( III), and both exit foramina lie within a shallow fossa on the orbitosphenoidlaterosphenoid suture, as in Daspletosaurus ( Russell, 1970) . The trochlear appears thinner in FMNH PR2081 than it does in the endocast studied by Osborn (1912). All other cranial nerves exit along the ventrolateral margin of the endocast. The trigeminal ( V) is a large nerve that diverges into two major branches— the anterior ophthalmic and posterior maxillomandibularshortly after leaving the brain, although this is apparent only on the right side (see below). The abducens nerve ( VI) has not been reported from tyrannosaurid endocasts, but a very slender lateral structure is visible on the right side of the digital endocast, immediately ventral to the ophthalmic nerve ( Fig. 1 View FIGURE 1 ), which I interpret as part of the abducens pathway, although it is disturbing that no obvious connection can be seen between this channel and the pituitary fossa. The glossopharyngeal ( IX), vagus ( X), and accessory ( XI) branch off together through the metotic fissure, visible on the endocast as a thick wall posterior to the otic capsule. The canal for the hypoglossal nerve ( XII) is also large and is visible toward the back of the endocast.
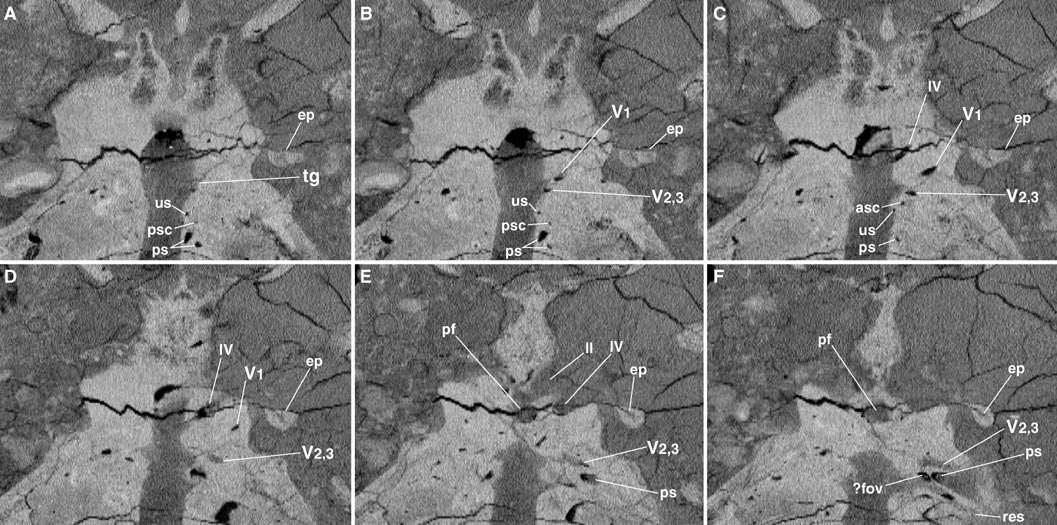
In hemisected tyrannosaurid braincases ( Osborn, 1912; Maleev, 1974) the facial ( VII) and both branches of the vestibulocochlear ( VIII) nerves exit through three very small foramina. These are visible in Osborn’s endocast as tiny nubs. Exits for the facial and the vestibular branch of the vestibulocochlear nerves are typically located immediately posteroventral to the trigeminal ganglion and directly ventral to the floccular recess. On the right side, a very slender channel can be seen directly ventral to the trigeminal nerves, but this is not likely to be the facial, as it occurs between V and VI. The external foramen for VII can be seen on the right side, both on the specimen and in the CT scans, but no connection can be made between it and this process on the endocast. On the left, two slender processes pass laterally and merge to form a dorsoventrally deep passage ( Fig. 2 View FIGURE 2 ). Its position is somewhat ventral to that of V on the right, and it clearly forms from two smaller channels. Moreover, there is a bulge on the endocast surface anterodorsal to them. The bulge may represent the trigeminal ganglion, with the discrete branches of V filled with dense matrix distally, and the smaller channels that merge may be the roots of the facial and vestibular nerves, which cannot be seen on the right. Based on this, ramification of the facial nerve into palatine and hyomandibular branches occurred outside the braincase. The foramen for the acoustic branch of VIII is also very small in theropods ( Osborn, 1912; Currie and Zhao, 1993; Makovicky and Norell, 1998); a slender process can be seen on the endocast medial to the semicircular canals, particularly on the left side, which may be either the acoustic nerve or the endolymphatic duct.
The opening identified by Osborn (1912) as the fenestra ovalis is, in fact, the caudal tympanic recess. Although not visible in the horizontal slices, the right stapes is preserved in FMNH PR2081 ; it passes through a crescentic, slitlike external otic recess ventral to the caudal tympanic recess ( Fig. 3F View FIGURE 3 ). We cannot unambiguously see the fenestra ovalis or fenestra pseudorotunda, though what I infer to be the fenestra ovalis is indicated in Figure 3F View FIGURE 3 .
Branches of Trigeminal Nerve —The ophthalmic and maxillarymandibular branches of the trigeminal nerve diverge before they leave the endocranial cavity, but they do not diverge very far in physical endocasts. Furthermore, the trigeminal nerve divides outside the braincase in living crocodylians. For these reasons, a pair of lateral openings at the prootic–laterosphenoid contact have sometimes been interpreted as the exit foramina for the ophthalmic and maxillarymandibular branches of the trigeminal nerve (e.g., Osborn, 1912:fig. 7). But in the CT images, one can see the two major branches of the trigeminal diverging broadly within the prootic—the ophthalmic passing anteroventrolaterally into the laterosphenoid, and the maxillarymandibular passing ventrolaterally ( Fig. 3 View FIGURE 3 ).
The ophthalmic foramen is within the laterosphenoid and lies immediately anteromedial to the epipterygoid. A similar placement for this foramen has been recognized for at least some derived theropods ( Colbert and Russell, 1969; Currie and Zhao, 1993; Clark et al., 1994), including tyrannosaurids ( Russell, 1970; Kurzanov, 1976; Molnar, 1991). In this regard, the tyrannosaurid braincase is birdlike: the avian ophthalmic nerve also diverges broadly from the maxillarymandibular nerve before leaving the braincase, exiting either through a small foramen in the orbitosphenoid or through the optic foramen ( BubienWaluszewska, 1981).
From the literature, it is unclear how broadly this character is distributed. The ophthalmic branch diverges broadly from the maxillarymandibular in an Allosaurus endocast ( Madsen, 1976; Rogers, 1998, 1999), but how this relates to external foramina is unknown. Other theropod braincases are described with a single foramen ovale (e.g., Stovall and Langston, 1950; Kurzanov, 1976), and other theropod endocasts are not as clear in this regard (e.g., Gilmore, 1920:pl. 36). A single foramen ovale is usually figured for sauropodomorphs (e.g., Holland, 1924; Galton, 1990; Madsen et al., 1995) and ornithischians, sometimes with mention of a sulcus on the laterosphenoid for the ophthalmic branch (e.g., Ostrom, 1961; Norman, 1986; Dodson and Currie, 1990; Sereno, 1991; Sullivan and Williamson, 1999), although at least one ceratopsian endocast shows a broad internal divergence between the ophthalmic and maxillarymandibular branches ( Brown, 1914). Moreover, it is not always clear if the opening labelled ‘‘V’’ in a plate is the opening or an opening for the trigeminal.
If the ophthalmic foramen is widely separated from its maxillarymandibular counterpart, why are there two closelyspaced foramina on the prooticlaterosphenoid suture? CT images reveal the presence of a large sinus that expands ventrally to merge with the channel for the maxillarymandibular nerves ( Fig. 3 View FIGURE 3 ). The anteriormost of these two foramina is for the maxillarymandibular nerves, and the posteriormost opens into the sinus. This sinus may be pneumatic in nature, and it is confluent anteroventrally with the tympanic cavity. Carr and Williamson (1999) described the prootic of a new species of Daspletosaurus as ‘‘inflated’’; this may relate to the sinus seen in the prootic of FMNH PR2081 .
Olfactory Bulbs —It was obvious to Osborn—and to later observers ( Maleev, 1965; Molnar, 1991; Horner and Lessem, 1993)—that the tyrannosaurid olfactory tract was very large compared with other theropods, judging from the large diameter of its canal in the orbitosphenoids and sphenethmoid. But the braincase Osborn hemisected does not preserve the frontals, and so the endocast he described lacked the olfactory bulbs. The impressions for the olfactory tracts and bulbs are large, but incompletely preserved, in a partial endocranial mold made from the frontals and parietals of a small tyrannosaurid from the Hell Creek Formation ( Molnar, 1978) considered by some to represent Aublysodon (Molnar and Carpenter, 1989) , but which may have been a very immature animal at the time of death. The bulbs are preserved in FMNH PR2081 as a pair of expansions from the tract anterior to the orbits, and they are extremely large ( Fig. 1 View FIGURE 1 ). The ventral margins of the bulbs in Figure 1 View FIGURE 1 are approximate, as there is no ossified floor for these structures in dinosaurs, but their dorsal and lateral extent is welldefined by the frontals and lacrymals. The bulbs are incomplete on the digital endocast, but as seen in a horizontal slice taken through the endocranial cavity ( Fig. 4B View FIGURE 4 ), they extend far anteriorly. Each bulb is approximately 1.5 times as wide as the cerebral region of the endocast ( Fig. 4 View FIGURE 4 ). There is no impression at the front of the olfactory fossa for an accessory bulb, suggesting that like living archosaurs ( Negus, 1958; Kuhlenbeck, 1977; Butler and Hodos, 1996), Tyrannosaurus lacked a vomeronasal organ.
The olfactory bulbs are smaller relative to brain size in other dinosaurs for which endocast information is available ( Osborn, 1912; Edinger, 1926; Russell, 1969; Currie, 1985, Rogers, 1998). Olfactory bulb size has been tentatively correlated with olfactory acuity in mammals and birds ( Smith, 1928; Papez, 1929; Cobb, 1960a, b; Bang and Cobb, 1968; Pearson, 1972; Bang and Wenzel, 1985; Butler and Hodos, 1996). This is true even for closely related birds of similar size; among North American cathartid vultures, the smelloriented turkey vulture has a larger olfactory bulb than does the visuallyoriented black vulture ( Bang, 1964). The large size of the bulbs in FMNH PR2081 is thus congruent with earlier suggestions that tyrannosaurids placed strong emphasis on their sense of smell.
It is premature to draw ecological conclusions from this observation. Although some variation in olfactory bulb size in birds may be correlated with food finding, other behaviors, such as mate location, have also been implicated ( Cobb, 1960a, b; Bang and Cobb, 1968). Moreover, until detailed information about the relative size of the olfactory bulbs for other gigantic theropods (e.g., Carcharodontosaurus , Giganotosaurus ) is available, we cannot rule out an allometric explanation for the large size of the olfactory bulbs in T. rex .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |