
Vareuptychia Forster, 1964
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4858.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4CDBAC51-FBCA-4157-B361-F56CA3EAC107 |
|
DOI |
https://doi.org/10.5281/zenodo.4498509 |
|
persistent identifier |
https://treatment.plazi.org/id/D33D878E-FFB6-7462-FF33-FA815450F8A5 |
|
treatment provided by |
Plazi |
|
scientific name |
Vareuptychia Forster, 1964 |
| status |
|
Vareuptychia Forster, 1964 View in CoL stat. rest.
Vareuptychia Forster, 1964: 125 View in CoL , nota.
Type species. Euptychia similis Butler, 1867 by original designation.
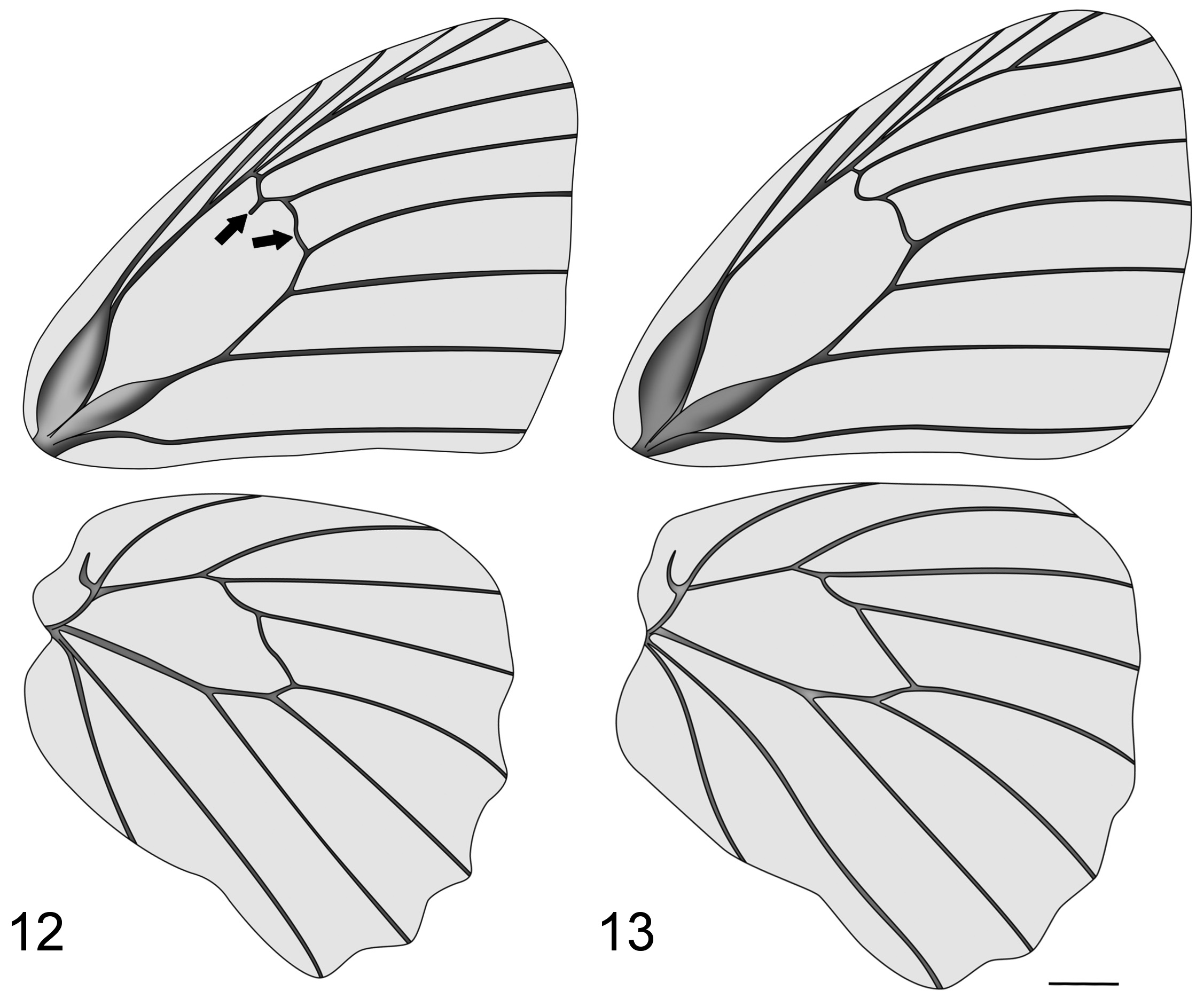
Diagnosis. Among the species of Euptychiina , the dorsal wing patterns of Vareuptychia stat. rest. species most resemble those of Yphthimoides Forster, 1964 (e.g. Y. manasses (C. Felder & R. Felder, 1867) , Y. renata (Stoll, 1780) and Y. affinis (Butler, 1867)) by the developed ocellus on CuA 1 -CuA 2 on the DHW. However, the VHW of Vareuptychia species are distinct from Yphthimoides , being more similar to some species of Moneuptychia Forster, 1964 (e.g. M. montana Freitas, 2015 and M. pervagata Freitas, Siewert & Mielke, 2015 ) in the shape and location of the ocelli in M 2 -CuA 1. Vareuptychia species differ from the above-mentioned Moneuptychia species by having four small ocelli in R 4+5 -M 1 (ocellus 1), M 1 -M 2 (ocellus 2), M 2 -M 3 (ocellus 3) and M 3 -CuA 1 (ocellus 4) on the VFW (sometimes the latter is not evident in some individuals), with ocellus 3 displaced towards the median line (not displaced in Moneuptychia ), vein dci concave and projected into the discal cell on the FW ( Fig. 12 View FIGURES 12–13 ), and the gnathos developed (absent in Moneuptychia ).
Redescription. Antennae half-length of costal margin of FW, with white scales around base of each flagellomere; chaetosema brown; postgenal area with small cream scales; densely hairy eyes; labial palp about two times length of head with small dark brown scales on dorsal and ventral sides of distal segment, small cream scales in lateral view of entire palp, and mixed elongated dark brown and cream scales at mid and basal segments. Legs covered with mixed small cream and brown scales with a paired tibial spur.
Wings. FW sub-triangular, costa slightly convex, apex rounded, external margin slightly convex without indentations, and inner margin straight. Ground color brown with three narrow dark brown lines (median, submarginal and marginal lines) on both DW, and a large ocellus in CuA 1 -CuA 2 with a black ocellar spot surrounded by a yellowish ocellar ring and two tiny silver-bluish pupils (some individuals can have only one pupil) on DHW. VFW with four dark brown or rufous narrow lines (median, submedian, marginal and submarginal), space between median and submarginal lines lighter with a dark umbra extending from costa to 2A, three to four small ocelli between R-M 1, M 1 –M 2, M 2 –M 3, M 3 –CuA 1, submarginal and marginal lines well-crenulated. HW sub-oval, somewhat longer than wide, costa convex, apex rounded, external margin convex with indentations, inner margin slightly sinuous. VHW with five ocelli from Rs-CuA 2, ocelli in Rs-M 1, M 1 -M 2 and CuA 1 -CuA 2 with black ocellar spot surrounded by yellowish ocellar ring (presence of pupils variable), ocelli in M 2 -CuA 2 with lighter ocellar spot and large silver pupils inside, ocellus in M 2 -CuA 1 elliptical and displaced towards median line. Some individuals can have an additional tiny ocellus in 2A and inner margin.
Venation. FW with veins Sc and CuA strongly swollen at base, 2A slightly swollen and sinuous at base; discal cell half-length of wing with a reduced recurrent vein and m 2 -m 3 sinuous. HW with discal cell half-length of wing, humeral vein developed and m 1 -m 2 convex.
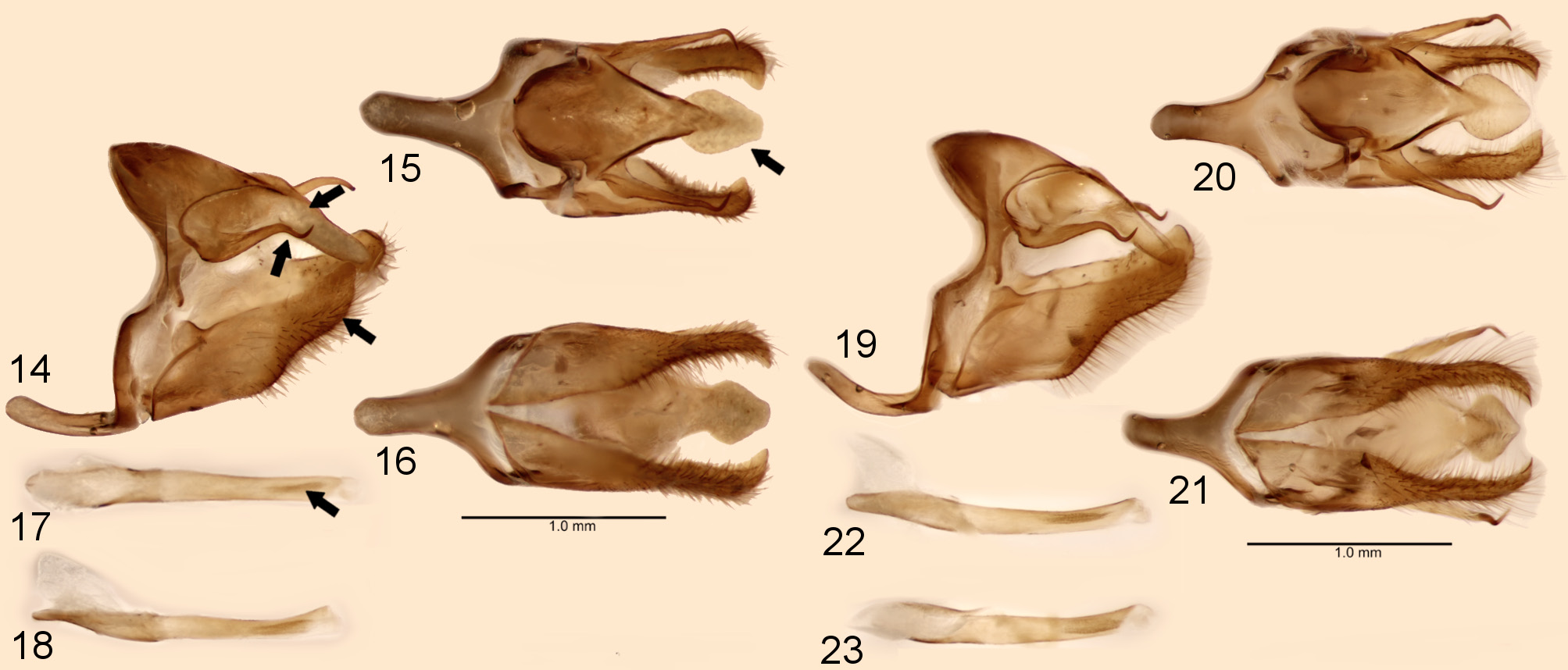
Male genitalia. Tegumen flattened in dorsal view, laterally subtriangular. Uncus robust, almost same length as tegumen, dorsally with mid-apical region ovoid and apex truncated. Gnathos sinuous laterally, half-length of uncus, with apical region hooked. Combination of ventral arms of tegumen and dorsal arms of saccus sinuous. Appendices angulares short. Anterior projection of saccus developed and cylindrical. Fultura superior absent. Fultura inferior a thin sclerotized stripe. Valva rhomboid covered by long hair-like setae latero-ventrally, and short setae at inner side, costa developed and sub-squared, dorsal margin straight, apical projection pointed and longer than apex of uncus, ventral margin slightly projected at median region. Aedeagus slightly curved upwards, short than valva, anterior region bottle-shaped, posterior region about 1.5 times longer than anterior region, distal margin dorsally truncated, distal opening ventral smaller than proximal opening. Vesica with two parallel cornutal patches ( Fig. 17 View FIGURES 14–23 ).
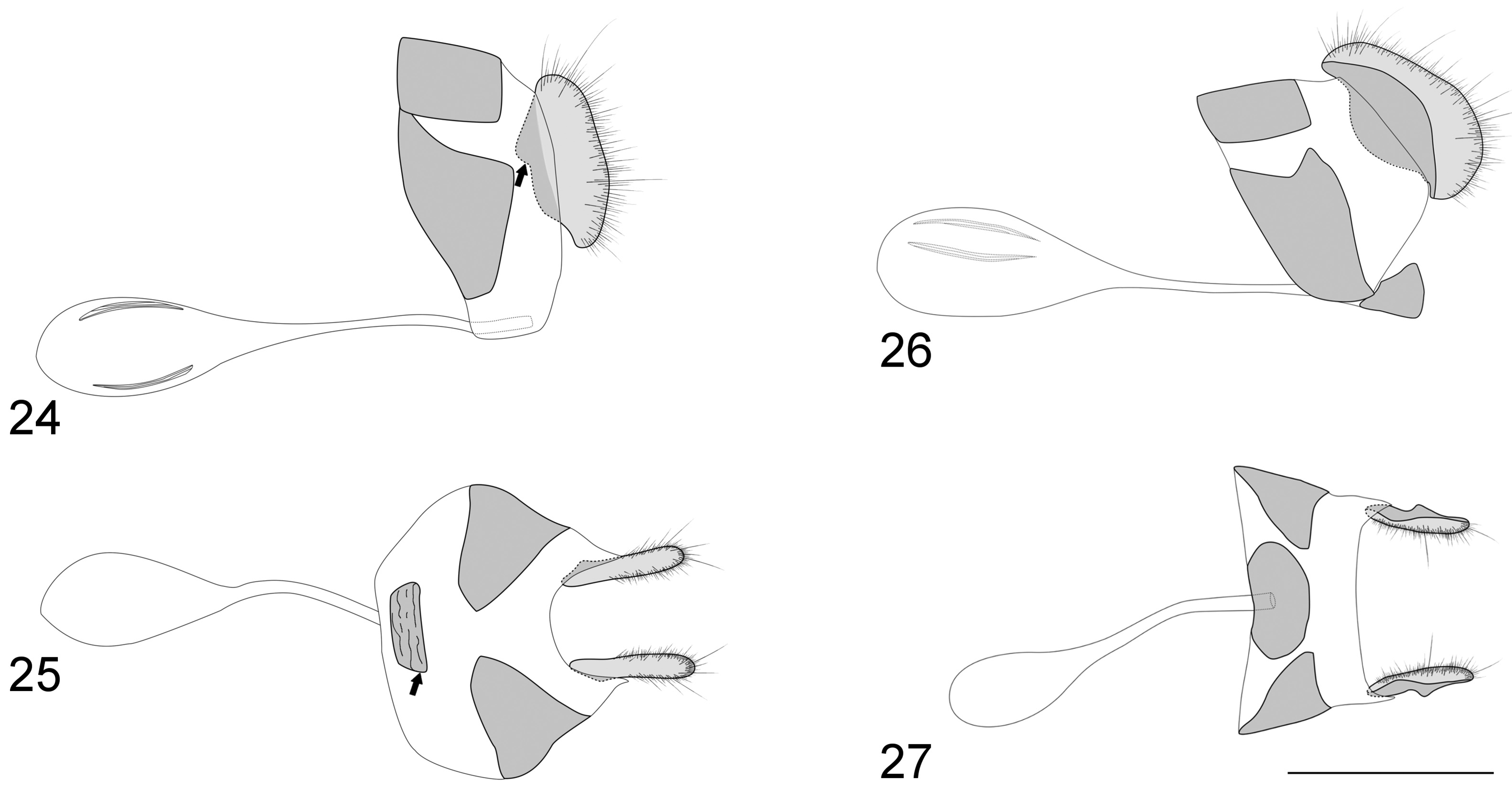
Female genitalia. Eighth tergite rectangular ( Fig. 24 View FIGURES 24–27 ). Papilla anales somewhat oblong, covered by long hairlike setae at distal region; posterior apophysis absent or reduced. Lateral plate (probably derived from 8 th sternite) can reach or not 8 th tergite and lamella antevaginalis. Lamella antevaginalis rectangular or half-orbicular in ventral view. Lamella postvaginalis absent. Ductus bursae membranous; corpus bursae smaller than ductus bursae, with paired signa of variable location.
Systematic position and discussion. Vareuptychia was erected by Forster (1964) to include Euptychia similis Butler, 1867 (the type species) and Euptychia usitata Butler, 1867 . In that work, Forster provided a description comparing Vareuptychia with Argyreuptychia Forster, 1964 (currently a subjective synonym of Cissia— see Zacca et al. 2018b) based on the wing color and shape of the male genitalia, although he did not specify which structures should be used to recognize the species of Vareuptychia . The male genitalia illustrations of V. similis and V. usitata in Forster (1964: 125 , figures 145 and 146, respectively) are quite different, and the genitalia of V. usitata agrees with Cissia pompilia (C. Felder & R. Felder, 1867) supporting its synonymy with the latter name ( Lamas 2004; Zacca et al. 2018b).
Lamas (2004) treated Vareuptychia as a subjective synonym of Cissia , and several authors accepted this proposal after him. However, based on morphological ( Marín et al. 2017) and molecular ( Murray & Prowell 2005) phylogenies, and the phylogenetic analysis reported here ( Fig. 87 View FIGURE 87 ), Vareuptychia is not closely related to the type species of Cissia , and morphological features from the wing pattern, venation, and male and female genitalia support recognition of Vareuptychia as a distinct genus. Compared to Cissia species (see Zacca et al. 2018b), Vareuptychia species can be distinguished by hairy eyes (glabrous in Cissia ), FW discal cell half length of wing (2/3 length of the wing in Cissia ), recurrent vein present in males (absent in Cissia ) and m 2 -m 3 sinuous (straight in Cissia ) ( Fig. 12 View FIGURES 12–13 ), robust uncus (slender in Cissia ), truncated apex of aedeagus (bifid apex in Cissia ) ( Fig. 17 View FIGURES 14–23 ), vesica with two cornutal patches (cornuti absent in Cissia ) ( Fig. 17 View FIGURES 14–23 ) and lamella antevaginalis with a wrinkled appearance and half-orbicular in ventral view (orbicular or obovate in Cissia ) ( Fig. 25 View FIGURES 24–27 ). Miller (1968) cited the tibial spurs on the midlegs as a probable synapomorphy of Cissia . However, such tibial spurs are also found in Vareuptychia species, and also frequently found in other euptychines (e.g. Magneuptychia , Graphita Nakahara, Marín & Barbosa, 2016 ), although their presence is variable in some genera, such as Euptychia Hübner, 1818 (S. Nakahara, pers. obs.). The male genitalia morphology is somewhat homogeneous in Vareuptychia stat. rest. species while the female genitalia have more informative characters to distinguish both species.
In the present study, Vareuptychia stat. rest. was recovered as monophyletic with high support (aLRT = 100 /BS = 100), but its sister relationship with Forsterinaria Gray, 1973 was moderately supported (aLRT = 91.3/BS = 76). In a more comprehensive phylogenetic study including>300 species of Euptychiina (Espeland et al., in prep.) Vareuptychia stat. rest. was recovered as sister to a clade containing some species of “ Magneuptychia ” (i.e. “ M”. analis (Godman, 1905) , “M”. modesta (Butler, 1867)) that will be allocated to a new genus (Nakahara et al., in prep.).
The ML analysis did not recover V. similis comb. rest. and V. themis comb. nov., as identified based on morphological characters, as reciprocally monophyletic ( Fig. 87 View FIGURE 87 ), which might suggest that the morphological characters actually represent intraspecific variation. Alternatively, this result might be an artifact of the low number of specimens used for DNA analysis, or due to incomplete lineage sorting as a result of a recent speciation event (Freeland 2006). We chose a more conservative approach and considered V. similis comb. rest. and V. themis comb. nov. as distinct species rather than as variations of a single species based on consistent differences in the wing phenotype that are not related to seasonal variation or sexual dichromatism (i.e. presence/absence of pupils in the ocellus at CuA 1 -CuA 2 in VHW), and the morphology of the female genitalia (i.e. lamella antevaginalis), in addition to the fact that these morphologically-defined species are sympatric and syntopic in some localities in Mexico and Guatemala with no intermediate phenotypes (see Examined material in Suppl. Material—S2; E. Pfeiler pers. comm.). Furthermore, numerous authors with first-hand experience of these butterflies in the field have continued to regard them as representing two species (see references listed below under each species). Nevertheless, future studies should focus on obtaining more DNA samples of Vareuptychia species throughout their entire distributional range ( Mexico to Colombia—Trinidad and Tobago), and population genetic approaches, to try to better elucidate the real diversity within the genus.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Satyrinae |
Vareuptychia Forster, 1964
| Zacca, Thamara, Casagrande, Mirna M., Mielke, Olaf H. H., Huertas, Blanca, Espeland, Marianne, Freitas, André V. L., Willmott, Keith R., Nakahara, Shinichi & Lamas, Gerardo 2020 |
Vareuptychia
| Forster, W. 1964: 125 |