
Diparinae, Thomson, 1876
|
publication ID |
https://doi.org/10.11646/zootaxa.1647.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9CDBECB7-17F1-4B0B-B577-CE29B34AA89A |
|
persistent identifier |
https://treatment.plazi.org/id/D40DA74B-DE50-5464-AE8F-63D8FBDEB948 |
|
treatment provided by |
Felipe |
|
scientific name |
Diparinae |
| status |
|
Phylogenetics of Diparinae
Taxonomic scope
A preliminary examination of the genera of Diparinae was conducted in order to remove taxa which obviously belonged to other groups or which could not be coded for a sufficient number of characters. A large number of diparine genera were described in the early twentieth century, when classification of Chalcidoidea was still in its early stages. Since Diparinae has never been revised on a world level, many genera traditionally classified within the group have obvious affinities to other subfamilies but have never been removed from Diparinae . These taxa were removed prior to the phylogenetic analysis because many of the characters used in the matrix had little relevance to these taxa. Taxa which were removed from the analysis because of insufficient coding were generally known only from males (making it impossible to code them for the large number of female-only characters), or in one case the type material was not located.
Specifically, Bekilliela Risbec and Dinarmolaelaps Masi were excluded from the analysis because neither are diparines (their taxonomic placement is discussed in the sections at the end of the paper on taxa removed or removed and placed in synonymy), and both genera are known only from males. Seyrigina Risbec was excluded from the analysis because it belongs in Eulophidae and is known only from the holotype, of which both the head and gaster are missing. Diparomorpha Hedqvist does belong in Diparinae , but was excluded from the analysis because it is known only from the holotype, which could not be located, and the description did not provide enough information to code the genus for a sufficient number of characters. The placement of Diparomorpha is discussed further in the generic entry for Dipara . The four Hawaiian genera ( Calolelaps Timberlake , Mesolelaps Ashmead , Neolelaps Ashmead , and Stictolelaps Timberlake ) were coded in the matrix but excluded from the analysis, because after coding they were determined to belong in Miscogasterinae and also to reduce the taxon:character ratio in the phylogenetic analysis.
For the most part, taxonomic units in the data matrix were genera. Exceptions to this were clades of genera in which a large number of undescribed species did not fit into the historical generic concepts ( Australolaelaps and Neapterolelaps ; Dipareta , Malinka , Nosodipara , and Pseudoceraphron ; and Conophorisca , Dolichodipara , and Myrmicolelaps ). New species were described to better understand phylogenetic relationships within those groups, and were included in the analysis. Dipara sensu Bouček (1988) was divided into multiple taxonomic units to better resolve its relationships with Alloterra , Grahamisia , Parurios , and Pseudipara . These divisions were based on groups which could be coded relatively unequivocally in the data matrix, as the variation within Dipara sensu Bouček (1988) made the coding of too many characters ambiguous. Additionally, separating Dipara sensu Bouček (1988) from Parurios on a world level proved impossible given the definitions of both genera. Additional species of Dipara were not described due to both time constraints and the sheer number of species which would need to be described to thoroughly examine this group. The subdivisions of Dipara sensu Bouček (1988) for the phylogenetic analysis include:
• Dipara turneri Hedqvist , a species from Africa which appears to lie between Dipara and Parurios mor-
phologically;
• <Fijian Dipara / Parurios >, an unclassifiable species from Fiji which shares some features with Dipara
turneri;
• <Indonesian Pondia >, a group of undescribed, Dipara -like species known from Indonesia and Taiwan,
which are often identified as Pondia in museum collections due to their convex scutellum;
• <Australian Dipara >, Girault’s Australian genera ( Epilelaps and Pseudiparella ) which Bouček (1988)
synonymized with Dipara ;
• <Micro Dipara >, generally minute, macropterous species of Dipara sensu Bouček (1988) with collar-
like pronota;
• Dipara sensu stricto, which are generally apterous, have a laterally bulging pronotum, and include the
most well-known members of the genus (e.g., D. canadensis ).
Outgroups selected included all three genera of Ceinae ( Bohpa , Cea , Spalangiopelta ), and two representatives each from Eunotinae ( Eunotus , Moranila ) and Coelocybinae ( Lelapsomorpha , Ormyromorpha ). Ceinae was thoroughly represented because Darling (1991a) proposed a sister-group relationship between Ceinae and Diparinae based on two morphological features: the presence of admarginal setae and papilliform antennal sensilla. Additionally, in a morphological study of Pteromalidae by Török and Abraham (2001), Ceinae grouped as sister to Diparinae in their reweighted parsimony tree with all taxa included. Bohpa Darling (Ceinae) was coded for all characters and analyses were run both including and excluding this taxa for three reasons. First, the male of Bohpa is unknown. With a wingless female and unknown male, it is impossible to code Bohpa for many of the characters important at the base of the phylogeny, including presence/absence of admarginal setae and the number of anelli in the male. Second, the addition of a third ceine genus does little to polarize characters within the clade, as among three taxa a gain and a loss are equally as parsimonious as two independent gains of a character state. Third, the inclusion of Bohpa in the analysis dramatically influenced the successive weighting of characters important to the phylogeny the derived diparines, which is discussed below. Eunotinae and Coelocybinae were chosen as additional outgroups based on discussions with other chalcidologists about which subfamilies might be closely related to Diparinae .
Character coding
All taxa were coded for 76 morphological characters. When possible, all characters, unless noted as “male”, were taken from female specimens. If a female specimen was unavailable, or the character of interest was damaged on all female specimens but visible on a male, the character was coded from the male. Exceptions to this are noted as (female), in which case sexual dimorphism causes females to have distinctive morphology from males for this character. Taxa coded with numbers in bold are polymorphic for that character, i.e. different species in the taxonomic unit have distinctly different states. Taxa coded with numbers in bold italics are ambiguous for that character, i.e. the state present in the taxon is intermediate between multiple coded states. Coding for all taxa is listed in Appendix I.
One of the features typically used to diagnose Diparinae is the presence of strong, dark bristles on the vertex and dorsal surface of the mesosoma. During the course of character coding for this analysis, it was noticed that although many diparines have a varying numbers of bristles, they always had bristles positionally identical to a subset of the full complement of bristles. To utilize this positional homology information in the phylogenetic analysis, bristles were coded in the following way: First, a single character for overall presence or absence of bristles was coded for all taxa (i.e., every taxon with at least 1 pair of bristles was coded as present, #21). Second, an additional presence/absence character was coded for each positionally homologous pair of bristles (#22–24, 26–29), as was a final additional character on the position of the median scutal bristle pair (#25). All bristle positional characters were coded as missing (?) if overall bristle presence/absence was coded as absent. This was intended not to give a disproportionate amount of weight to the presence/absence of bristles, as it is plausible that all bristles could be lost in a single evolutionary step.
Two issues arise from this coding scheme, however. First, by coding positional bristle characters for all taxa without an overall presence/absence character, the large number of bristle characters themselves could drive much of the phylogenetic signal in the tree if they supported the same groupings. Second, strange step requirements can occur when reconstructing character states. For example, a taxon without bristles can be reconstructed as sister-group to a taxon with all bristle pairs with a single step (overall bristle presence/ absence). To then place a taxon with a single bristle pair within that clade as sister to the bristleless taxon would require at least 6 additional steps, while this transformation seemingly should only take 1 step. A character could be constructed in which all possible bristle conformations are coded as different states (which was done with notaular shape), but the vast number of potential states would prove uninformative in the analysis. Therefore, analyses were run both including and excluding positional bristle characters.
Additionally, the form of the notauli was originally coded as multiple characters (#32–35). However, notauli appear to be highly divergent in closely related taxa, and having multiple characters would imply a heavy weighting. In order to minimize the effect of notauli on tree topology, characters #32–34 were combined into a single character representing all possible arrangements of the notauli (#31), and characters #32– 34 were excluded from all analyses. Character #35 (notaular pads) was left as is, because, although the black pads are located adjacent to the notauli, they are not part of the notaular structure.
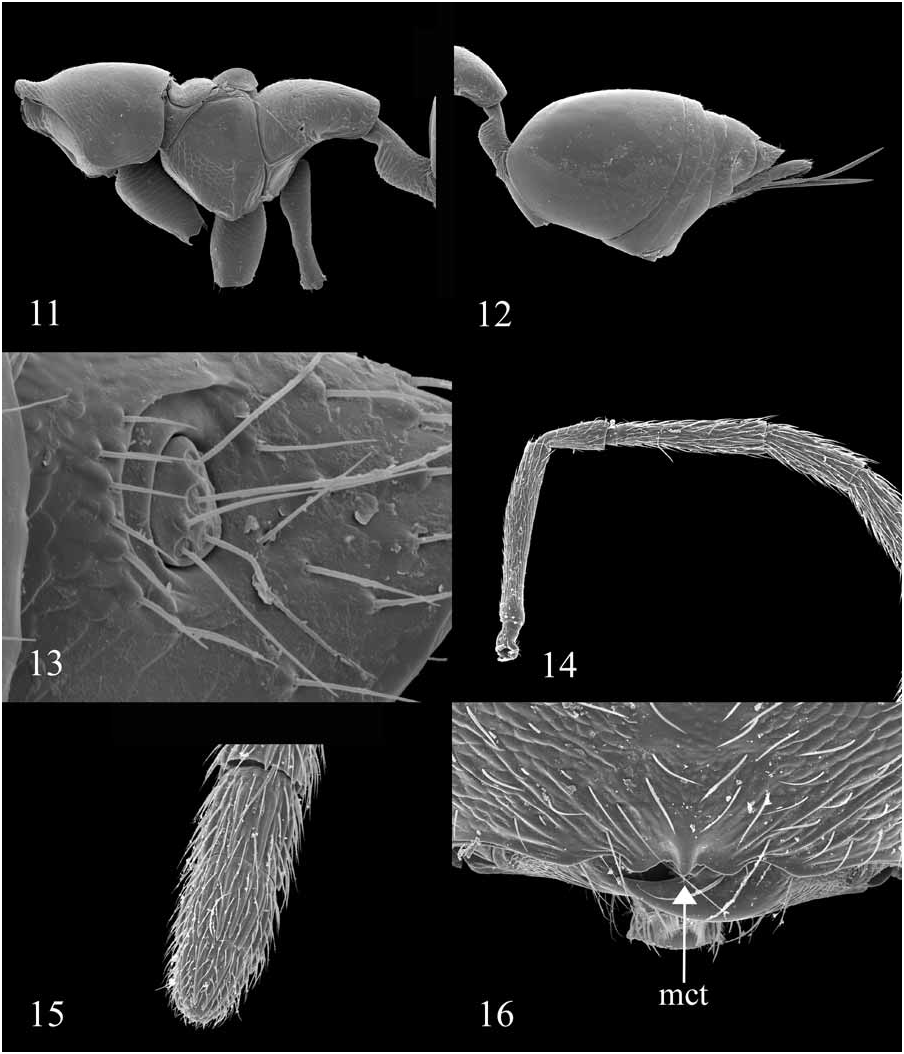
1. Apical Clypeal Margin: 0 = concave ( Fig. 45 View FIGURES 41–46 ); 1 = with median tooth or lobe ( Fig. 16 View FIGURES 11–16 , mct); 2 = protruding, symmetrically bilobed (lobes may be very small and appear symmetrically sinuate); 3 = convex, or protruding and straight.
A convex clypeal margin is coded the same as a protruding clypeus with a straight margin, as the blurring between these two forms makes them difficult to code as separate. Additionally, both lack the lobes or teeth of states 1 or 2, and project from the ventral margin of the head, as opposed to state 0. The clypeal margin has not been previously used in diparine taxonomy, with the exception of Lelaps and Spalangiolaelaps , which have often been partially diagnosed by a median tooth (e.g., Yoshimoto 1977). In his phylogenetic analysis of Toryminae, Grissell (1995) used a more finely detailed transformation series of the clypeal apex, which was highly consistent within the phylogeny (CI = 1.00).
2. Posterior Margin of Gena: 0 = smooth; 1 = carinate.
Outside of Diparinae , a carinate genal margin is found sporadically in Chalcididae ( Wijesekara 1997) , Eucharitidae ( Heraty 2002) , and also in Eurytomidae and New World Lyscini ( Pteromalidae : Cleonyminae ) ( Gibson 2003). Gibson (2003) states that this feature is often associated “with a head that is closely appressed to the pronotum so the two form a more rigid association.”
3. Dorsal Margin of Scrobe: 0 = rounded; 1 = carinate.
This character is sporadically distributed throughout Diparinae . The presence of a carinate dorsal margin is probably correlated with the overall “carinateness” of a chalcidoid, i.e. diparines which are heavily sculptured often have carinate margins of many features.
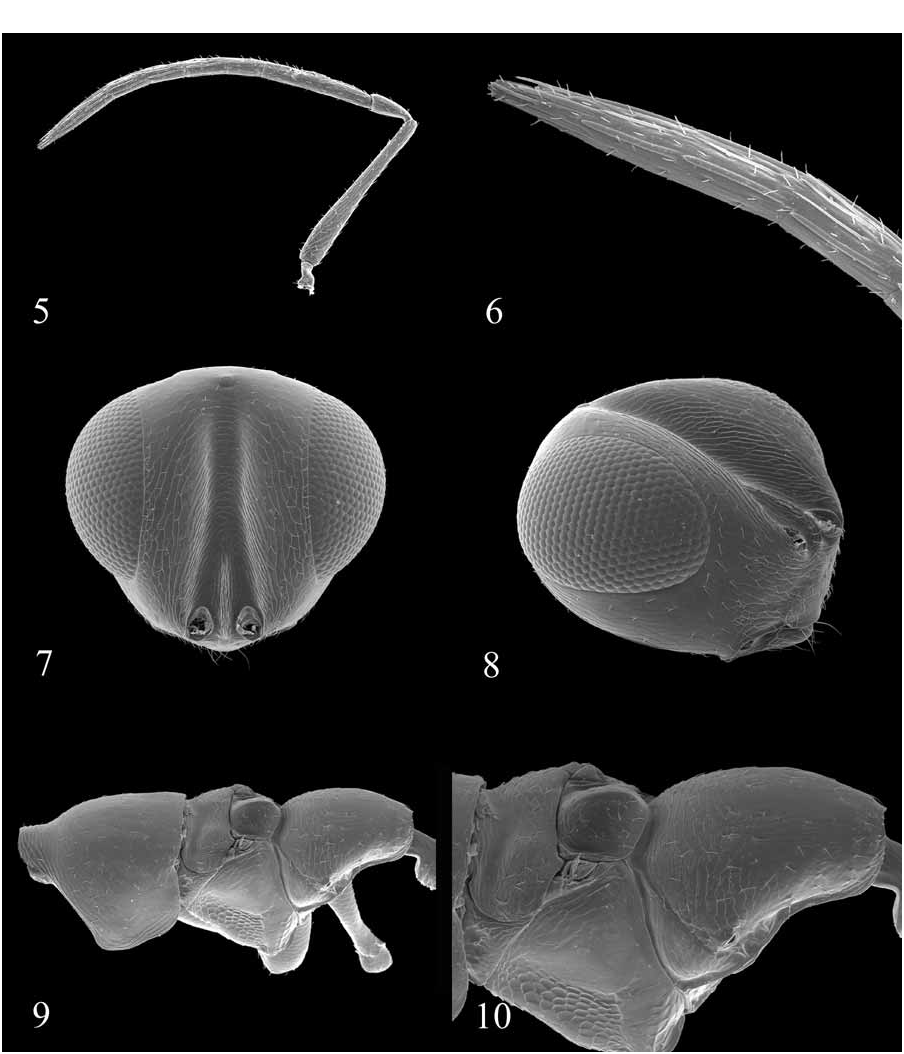
4. Scrobe Shape: 0 = scrobe present and scrobal channel parallel-sided ( Figs. 7 View FIGURES 5–10 , 27 View FIGURES 23–28 ); 1 = scrobe present and scrobal channel wide, triangular dorsal to toruli ( Fig. 36 View FIGURES 35–40 ); 2 = wide, shallow depression, scrobe basically absent ( Fig. 44 View FIGURES 41–46 ); 3 = scrobe distinct and short, but without scrobal channel ( Fig. 52 View FIGURES 47–52 ); 4 = scrobe triangular and wide, but without scrobal channel.
State 3 is autapomorphic for Dipara < Pondia >. Gibson coded a similar character in his cleonymine (2003) and eupelmid (1989) phylogenetic analyses. In both groups he found that most members possessed scrobes with distinct scrobal channels (equivalent to state 0 in this analysis), although the scrobes had been reduced or lost in some members of both groups.
5. Occiptal Margin: 0 = rounded; 1 = carinate.
The occipital margin refers to the boundary between the posterior margin of the vertex and the dorsal margin of the occiput.
6. Occipital Carina: 0 = absent; 1 = present.
The presence of an occipital carina was initially coded as two separate states, one in which the carina was visible only as line dorsal to the occipital foramen, and another where the carina was a semicircular line, laterally reaching ventral to the occipital foramen. However, the ambiguity in coding this character for many taxa has led to the reduction of the presence of the occipital carina into a single state. Grissell (1995) hypothesized that the absence of an occipital carina is the ancestral state in Pteromalidae , as did Gibson (2003) regarding Cleonyminae .
7. Face Sculpture (female): 0 = Upper face without strong, transversely carinate sculpture ( Figs. 7 View FIGURES 5–10 , 27 View FIGURES 23–28 , 36 View FIGURES 35–40 , 44 View FIGURES 41–46 ); 1= Upper face with deep pits separated by strong transverse carinae.
This feature is unique to Dipara turneri and <Fijian Dipara / Parurios >, and was included to help discern the internal phylogeny of the Dipara clade. As the males of both these species are unknown, it is uncertain whether or not this sculpture is present in both sexes, and the character is therefore only coded from females.
8. Torular Shelf: 0 = toruli not on shelf, angle of 50° or less between upper face and lower face ( Fig. 20 View FIGURES 17–22 ); 1 = toruli on shelf, sharp or rounded angle of ~90° between upper and lower face ( Fig. 8 View FIGURES 5–10 ).
9. Malar Groove: 0 = present; 1 = absent.
The malar groove of Myrmicolelaps iridius is coded as?, because although there is no distinct line, a depressed area delimited by a change in sculpture marks the location of the malar groove. This character may not be independent from body size, as the malar groove tends to lost in the most diminutive taxa.
10. Posterior Eye Extension: 0 = normal, eye not extending posteriorly beyond occipital margin, pronotum visible in lateral view ( Fig. 20 View FIGURES 17–22 ); 1 = eye extended posteriorly beyond occipital margin, laterally obscuring pronotum.
This character is exists to a varying degree in the eunotines Eunotus and Moranila , and is coded as polymorphic for both taxa.
11. Inner Eye Margins: 0 = parallel-sided or uniformly convex; 1 = ventrally diverging.
This character has historically been used to help define the outgroup Coelocybinae (e.g., Bouček 1988) as well as Eupelmidae ( Gibson 1989) . Gibson (2003) found that diverging inner eye margins supported monophyly of Cleonyminae if the outgroup Hetreulophini ( Colotrechninae ) was excluded from the analysis and his interpretation of the character in Callimomoides was incorrect. However, Gibson did not include representatives of Eupelmidae and Coelocybinae in his analysis, which casts further doubt on the utility of this character for defining Cleonyminae . Regardless, due to the uniformity of distribution of this feature in pteromalid subfamilies and Eupelmidae , it may become an important character in the higher phylogeny of Chalcidoidea . Within the Diparinae , ventrally diverging eyes are unique to Cerodipara .
12. Eye Setae: 0 = eyes bare; 1 = eyes sparsely setose.
Eyes are only coded as setose if the setae are readily visible under microscopic examination. If the setae are so small and sparse as to only be visible at certain angles of light and high magnification, they are considered bare. Unforunately, a vast majority of diparines fall into this latter category, and the coding of this character may be heavily influenced by personal interpretation. Eye setation appears fairly variable within Diparinae , and shows a similar pattern of variability in both Cleonyminae ( Gibson 2003) and Eupelmidae ( Gibson 1989, 1995).
13. Antennal Shape (Female): 0 = filiform; 1 = clavate; 2 = clubbed.
14. Antennal Symmetry (Female): 0 = symmetrical; 1 = asymmetrical.
An asymmetrical flagellum is typically characteristic of Coelocybinae ( Bouček 1988) , although some New World coelocybines have a symmetrical flagellum ( Heydon and Hanson (2005). Within Diparinae , an asymmetrical flagellum is unique to Netomocera .
15. Anellar Number (Female): 0 = 1 anellus; 1 = 2 anelli; 2 = 3 anelli; 3 = 5 anelli; 4 = 7 anelli.
In cases where the anellus is partially fused to the first funicular segment (e.g. Myrmicolelaps aurantius , see Fig. 26 View FIGURES 23–28 ), these taxa are coded as having a single anellus.
16. Flagellar Number (Female): 0 = 11 flagellar segments (counting clava as 3); 1 = 8 or fewer flagellar segments (counting clava as 3).
A reduced number of flagellar segments is often used as a diagnostic feature of Eunotinae (e.g., Bouček 1988).
17. Pedicel:F1 (Female): 0 = first funicular segment subequal in length to pedicel; 1 = first funicular segment at least 1.5X as long as pedicel; 2 = pedicel at least 1.5X as long as first funicular segment.
Taxa between states (e.g., pedicel 1.3X as long as F1) were coded as ambiguous. Gibson (1997) used a version of this character “first funicular segment longer than pedicel” in their generic key for separating Lelaps and Spalangiolaelaps from other Nearctic diparines. However, this character does not separate Lelaps and Spalangiolaelaps from many other non-Nearctic diparines.
18. F1:F2 (Female): 0 = First funicular segment subequal in length to second funicular segment; 1 = First funicular segment at least 1.5X longer than second funicular segment.
All species of Pseudoceraphron are coded as N/A for this character, because they have 7 anelli and only a single funicular segment. Yoshimoto (1977) used an elongate F1 to define the tribe Lelapini ( Lelaps and Spalangiolaelaps ) in his key. Nosodipara ferrana also has F1 at least 1.5X longer than F2, although this is due to a shortened F2 rather than an elongate F1.
19. Claval Fusion (Female): 0 = clava 3-segmented ( Figs. 15 View FIGURES 11–16 , 43 View FIGURES 41–46 ); 1 = clava 2-segmented, distal 2 segments apparently fused; 2 = clava 1-segmented, all segments apparently fused ( Figs. 5, 6 View FIGURES 5–10 , 25 View FIGURES 23–28 ).
A two-segmented clava is autapomorphic for Conodipara . Gibson (2003) used a similar coding in his phylogenetic analysis of Cleonyminae . This character appears to show a moderate degree of variability in both the Cleonyminae ( Gibson 2003) and Eupelmidae ( Gibson 1989) .
20. Claval Micropilosity (Female): 0 = clava without thick tuft of micropilosity on apex of clava ( Figs. 5, 6 View FIGURES 5–10 , 15 View FIGURES 11–16 , 25 View FIGURES 23–28 , 36 View FIGURES 35–40 ); 1 = clava with thick tuft of micropilosity on apex of clava ( Figs. 42, 43 View FIGURES 41–46 ).
The micropilosity at the apex of the clava of Pseudoceraphron and Nosodipara appear to be sensilla rather than setae. In Figure 43 View FIGURES 41–46 , it can be seen that the dorsal sensilla appear to have closed, rounded tops, while the ventral sensilla appear to have open, truncated tops. It is possible that the tips of the ventral sensilla were sheered off during SEM preparation, although the uniformity of the open-topped sensilla suggests that is unlikely.
21. Bristles (Presence/Absence): 0 = without any strong, dark bristles on the thorax or vertex, excluding bristles emanating from the wing stump in apterous specimens; 1 = at least 1 pair of strong, dark bristles present on the thorax or vertex, excluding bristles emanating from the wing stump in apterous specimens.
Strong, dark bristles are used by many keys to diagnose Diparinae (e.g. Bouček 1988, Gibson 1997), although many diparine genera lack bristles completely.
22. Bristles (Vertex): 0 = absent; 1 = bristle pairs limited to occipital margin and dorsal inner eye margins ( Fig. 18 View FIGURES 17–22 ); 2 = bristles uniformly distributed across vertex; 3 = with only a single pair of bristles posterior to the lateral ocelli.
Not only do the bristle patterns differ fundamentally between states 1 and 2, but the bristles of Lelapsomorpha are significantly thinner than most taxa having state 1 (although this difference was too difficult to code, and may not be entirely independent from the pattern itself). State 3 is unique to Chimaerolelaps .
23. Bristles (Pronotal): 0 = without bristles on pronotum; 1 = with transverse row of bristles on pronotum; 2 = with a single pair of bristles on prontum.
24. Bristles (Medial Scutal): 0 = without pair of bristles on scutum medial to notauli; 1 = with a single pair of bristles on scutum medial to notauli; 2 = with two pairs of bristles on scutum medial to notauli.
State 2 is unique to Moranila .
25. Position of Median Scutal Bristles: 0 = normal, distance from scuto-scutellar margin to median scutal bristles longer than distance to anterior scutellar bristles; 1 = short, distance from scuto-scutellar margin to median scutellar bristles shorter than distance to anterior scutellar bristles.
Bouček (1988) used this character to separate female Dipara and Parurios in his key, although he stated that non-Australian Dipara would key to Parurios using this feature (therefore it would theoretically separate Australian Dipara from the remainder of the Dipara s. l. clade). This character is only coded in taxa which have both A) median scutal bristles, and B) a single pair of anterior scutellar bristles. All other taxa are coded as N/A.
26. Bristles (Lateral Scutal): 0 = without bristles on lateral lobes of scutum; 1 = with a single pair of bristles on the dorso-lateral margin of the lateral lobes of scutum ( Fig. 19 View FIGURES 17–22 ); 2= with a single pair of bristles on the antero-medial portion of the lateral lobes of scutum ( Fig. 22 View FIGURES 17–22 ); 3 = with two pairs of bristles on lateral lobes of scutum.
States 2 and 3 are possessed only by outgroup taxa. Moranila possesses a single pair of bristles which is not positionally homologous to those in the Diparinae , while Lelapsomorpha and some species of Spalangiopelta both possess two pairs of lateral scutal bristles.
27. Bristles (Anterior Scutellar): 0 = without pair of bristles on anterior half of scutellum (if frenal sulcus is absent) or anterior to frenal sulcus; 1 = with a single pair of bristles on anterior half of scutellum (if frenal sulcus is absent) or anterior to frenal sulcus; 2 = with two pairs of bristles on anterior half of scutellum (if frenal sulcus is absent) or anterior to frenal sulcus; 3 = with three pairs of bristles on anterior hald of scutellum (if frenal sulcus is absent) or anterior to frenal sulcus.
Two pairs of anterior scutellar bristles is unique to Lelapsomorpha , while three pairs is unique to Chimaerolelaps .
28. Bristles (Posterior Scutellum): 0 = without pair of bristles on posterior half of scutellum; 1 = with pair of bristles on posterior half of scutellum, either at posterior margin (if frenal sulcus is absent) or lateral margins of frenal sulcus; 2 = with pair of bristles on posterior half of scutellum, posterior to distinct frenal sulcus.
State 2 is autapomorphic for some species of Spalangiopelta .
29. Bristles (Propodeal): 0 = without pair of bristles on propodeum; 1 = with pair of bristles on lateral margins of propodeum.
Propodeal bristles are unique to Pseudoceraphron < Malinka >.
30. Pronotum Shape: 0 = short, collar-like, not dorsally or laterally bulging, and at least as wide as long ( Fig. 19, 20, 22 View FIGURES 17–22 , 46 View FIGURES 41–46 , 47, 48 View FIGURES 47–52 ); 1 = large, laterally but not dorsally convex; 2 = large, both laterally and dorsally convex ( Fig. 9 View FIGURES 5–10 ); 3 = large, cylindrical, longer than wide.
31. Notauli (combined): 0 = notauli absent; 1 = notauli present only in anterior half; 2 = notauli complete, Y-shaped; 3 = notauli strongly arched along entire length (appearing semi-circular) and meeting posterior scutal margin at scutoscutellar suture; 4 = notauli not arched and meeting posterior scutal margin exterior to or at the edge of scutoscutellar suture; 5 = notauli strongly arched anteriorly and running parallel posteriorly, meeting posterior scutal margin at scutoscutellar suture; 6 = notauli not arched and meeting posterior scutal margin at scutoscutellar suture; 7 = notauli inverted, U-shaped, meeting anterior to posterior scutal margin.
In all analyses shown here the combined notauli character (#31) was used rather than the subsequent separate notaular characters (#32–34) to avoid overweighting of notauli.
32. Notauli: 0 = notauli distinct along their entire length; 1 = notauli faded posteriorly; 2 = notauli absent.
Gibson (2003) hypothesizes that distinct notauli are pleisiomorphic for the Cleonyminae , although the character varies within the group and also in eupelmids ( Gibson 1989, 1995). Wijesekara (1997) coded all chalcidids as having distinct notauli, while they were completely absent in his leucospid outgroup. Grissell (1995) also hypothesized that distinct notauli were pleisiomorphic for Toryminae , although he also stated that the character was so homoplastic as to provide little information. Within the Diparinae only Pyramidophoriella lacks notauli completely.
33. Notaular Meeting: 0 = notauli posteriorly widely spaced, meeting posterior scutal margin exterior to or at the lateral edge of the scutoscutellar suture; 1 = notauli posteriorly narrowly spaced, meeting posterior scutal margin at scutoscutellar suture, or notauli meet at or just anterior to posterior scutal margin; 2 = notauli Y-shaped, meeting well anterior to posterior scutal margin.
Both Gibson (1989, 2003) and Grissell (1995) used similar characters. While Gibson (2003) found this character difficult to code for cleonymines, most eupelmids have notauli which are “distinctly exterior to the scutoscutellar suture” (= widely spaced in this analysis). Grissell (1995) coded all torymines as having widely spaced notauli except Palachia . Diparines with U-shaped notauli are coded as ambiguous for this character, as they often have extremely short scuta, making this difficult to code. Pseudoceraphron is coded as 0 for this character, as its notauli are widely spaced. The notauli do meet the posterior scutal margin within the boundaries of the scutellum, but this is because Pseudoceraphron has an extremely wide scutellum without axillae (and therefore no scutoscutellar suture).
34. Notaular Arch: 0 = notauli not arched, almost appearing as straight lines; 1 = notauli strongly arched along entire length, appearing semi-circular; 2 = notauli strongly arched anteriorly, but running parallel sided posteriorly; 3 = notauli U-shaped, arch inverted.
Diparines with Y-shaped notauli are coded as ambiguous for this character, as the short distance the notauli travel before meeting each other makes it difficult to code for this character.
35. Notaular Pads: 0 = lateral lobes of scutum without raised, black pads; 1 = lateral lobes of scutum with raised, black pads, lateral to the posterior central groove of Y-shaped notauli.
The presence of these notaular pads has been used to separate Dipara < Grahamisia > from Dipara sensu Bouček (1988) , and Pseudoceraphron < Malinka > from both Pseudoceraphron sensu Bouček (1988) and Pseudoceraphron < Dipareta >.
36. Posterior Scutal Margin: 0 = posterior scutal margin lateral to scutellum not grooved for the insertion of the anterior margin of the tegula ( Figs. 10 View FIGURES 5–10 , 19, 22 View FIGURES 17–22 , 38 View FIGURES 35–40 , 48 View FIGURES 47–52 , 55 View FIGURES 53–58 ); 1 = posterior scutal margin lateral to scutellum grooved for the insertion of the anterior margin of the tegula, with small cluster of setae on the medial edge of groove ( Fig. 30 View FIGURES 29–34 ).
37. Scutellum Shape (Female): 0 = large, convex, gently sloping down posteriorly ( Figs. 19, 22 View FIGURES 17–22 , 50, 51 View FIGURES 47–52 ); 1 = wider than long, sloping up and slightly pointed posteriorly ( Figs. 37, 38 View FIGURES 35–40 ); 2 = conical and tooth-like, narrowed anteriorly, slightly laterally compressed, and flattened posteriorly ( Figs. 9 View FIGURES 5–10 , 11 View FIGURES 11–16 , 54, 55 View FIGURES 53–58 ); 3 = small, strongly convex, not sloping posteriorly ( Fig. 53 View FIGURES 53–58 ); 4 = flat, wider than long ( Fig. 48 View FIGURES 47–52 ); 5 = square, slightly raised and laterally compressed medio-posteriorly ( Fig. 29 View FIGURES 29–34 ); 6 = large, slightly convex, not sloping posteriorly; 7 = slightly convex, wider than long, lateral and posterior margin with upturned carina (see Darling 1991b: Figs. 6, 9 View FIGURES 5–10 ).
There is a tremendous amount of morphological variation in the diparine scutellum. State 1, 6, and 7 appear to autapomorphic for Neapterolelaps , Dozodipara , and Bohpa , respectively. Dipara turneri is coded as state 0; although its scutellum is slightly flattened, it is still large and slightly convex.
38. Axillae (Female): 0 = normal, large and convex ( Figs. 19, 22 View FIGURES 17–22 , 50, 51 View FIGURES 47–52 ); 1 = reduced and convex ( Fig. 38 View FIGURES 35–40 ); 2 = reduced and concave or flat ( Figs. 10 View FIGURES 5–10 , 29 View FIGURES 29–34 ); 3 = entirely absent ( Fig. 48 View FIGURES 47–52 ); 4 = indistinct from scutellum, indicated only by faint sulcus between axilla and scutellum (see Darling 1991b: Figs. 1 View FIGURE 1 , 3 View FIGURE 3 ).
State 4 is autapomorphic for Bohpa .
39. Posterior Notal Wing Process (Female): 0 = present, pointed ( Figs. 19, 22 View FIGURES 17–22 , 50, 51 View FIGURES 47–52 ); 1 = absent ( Figs. 29 View FIGURES 29–34 , 46 View FIGURES 41–46 , 47 View FIGURES 47–52 ); 2 = present but squarely truncate ( Fig. 38 View FIGURES 35–40 ); 3 = present but truncate and rounded.
Many apterous diparines have a reduced or absent posterior notal wing process. This character may be an accurate measure of “potential” wing size, as taxa with both apterous and macropterous members have complete posterior notal wing processes even in their apterous members. State 2 is autapomorphic for Neapterolelaps .
40. Frenal sulcus: 0 = present; 1 = absent.
The frenum has been previously used to distinguish Lelaps < Spalangiolaelaps > (frenum absent) from the remainder of Lelaps (frenum present). Grissell (1995) used the presence/absence of the frenum in his phylogenetic analysis of the Toryminae and found it to be extremely homoplastic. Here the character is coded as presence/absence of the frenal sulcus, as the area of the scutellum homologous to the frenum itself is presumably still present. This character may not be entirely independent from scutellum shape, as taxa with conical scutella never possess a frenal sulcus.
41. Metanotum: 0 = normal, sculptured, narrow band ( Figs. 20, 22 View FIGURES 17–22 , 50, 51 View FIGURES 47–52 , 53 View FIGURES 53–58 ); 1= absent, or present only as smooth narrow strip ( Figs. 10 View FIGURES 5–10 , 38 View FIGURES 35–40 , 48 View FIGURES 47–52 ); 2 = smooth, high, and vertical.
42. Dorsellum: 0 = present as raised, convex medial region of metanotum; 1 = completely absent ( Figs. 10 View FIGURES 5–10 , 19 View FIGURES 17–22 , 29 View FIGURES 29–34 , 38 View FIGURES 35–40 , 48, 50, 51 View FIGURES 47–52 , 55 View FIGURES 53–58 ); 2 = modified into cup-like structure ( Fig. 22 View FIGURES 17–22 ).
Moranila (and the entire tribe of Moranilini) uniquely possesses a modified, cup-shaped dorsellum.
43. Propodeal Shape: 0 = propodeum normal, descending posteriorly in lateral view, but at least 1.5X longer than high ( Figs. 19 View FIGURES 17–22 , 37 View FIGURES 35–40 , 50, 51 View FIGURES 47–52 ); 1 = propodeum steeply descending posteriorly in lateral view, as long as high or higher than long; 2 = propodeum rising posteriorly from anterior margin for a least a portion of its length ( Figs. 9, 10 View FIGURES 5–10 , 54, 55 View FIGURES 53–58 ); propodeum dorsally flat in lateral view, wider than long in dorsal view ( Figs. 46 View FIGURES 41–46 , 47 View FIGURES 47–52 ); propodeum dorsally flat in lateral view, longer than wide in dorsal view ( Fig. 28 View FIGURES 23–28 ).
Propodeal height and length measurements are made from the median posterior margin of metanotum to the dorsal margin of propodeal foramen.
44. Propodeal Spine: 0 = without anterior or median spine ( Figs. 9, 10 View FIGURES 5–10 , 19, 22 View FIGURES 17–22 , 28 View FIGURES 23–28 , 29 View FIGURES 29–34 , 37, 38 View FIGURES 35–40 , 48, 50, 51 View FIGURES 47–52 , 55 View FIGURES 53–58 ); 1 = with anterior spine, sometimes with a median carina emanating posteriorly; 2 = with median spine with 4–6 carinae emanating from it.
This character is included to help in resolving the relationships between and within Dipara < Parurios > and Dipara sensu Bouček (1988) . Lelapsomorpha is coded as? for this character because it does have an anterior protrusion on its propodeum, however, the protrusion is wide and dorsally flat, and therefore difficult to code as a spine.
45. Plicae: 0 = absent, or present but propodeum not strongly depressed lateral to plicae; 1 = propodeum with longitudinal plicae, and propodeum strongly depressed lateral to plicae.
46. Postspiracular Area: 0 = normal, suture between postspiracular area and metapleuron diagonal ( Figs. 28 View FIGURES 23–28 , 37 View FIGURES 35–40 , 54, 55 View FIGURES 53–58 ); 1 = facing posteriorly, suture between postspiracular area and metapleuron vertical ( Fig. 49 View FIGURES 47–52 ).
47. Propodeal Foramen: 0 = normal, open in one plane ( Fig. 49 View FIGURES 47–52 ); 1 = hinge-like, open in two planes (posteriorly and ventrally) ( Fig. 31 View FIGURES 29–34 ).
The genera Conophorisca and Myrmicolelaps all possess a propodeal foramen which appears hinge-like (i.e. it appears as two parabolas abutting at right angles, opening both posteriorly and ventrally).
48. Prepectus Extension: 0 = reduced, not extending posteriorly to tegula ( Figs. 46 View FIGURES 41–46 , 47 View FIGURES 47–52 , 54 View FIGURES 53–58 ); 1 = elongate, extended posteriorly along lateral margin of scutum to anterior margin of tegula ( Figs. 20 View FIGURES 17–22 , 37 View FIGURES 35–40 ).
Conodipara , Conophorisca , and Myrmicolelaps all have a prepectus which comes close to but does not touch the tegula. All of these genera have extremely short scuta, and the reduced distance between the prepectus and the tegula is hypothesized to be the result of the scutal reduction rather than an extended prepectus. These genera are therefore coded as state 0.
49. Tegula Shape: 0 = normal, flap-like ( Figs. 20 View FIGURES 17–22 , 28 View FIGURES 23–28 , 30 View FIGURES 29–34 , 37 View FIGURES 35–40 ); 1 = extended anteriorly underneath lateral margin of scutum ( Figs. 46 View FIGURES 41–46 , 47 View FIGURES 47–52 , 54, 55 View FIGURES 53–58 ).
50. Longitudinal Invagination: 0 = lateral edge of pronotum and dorsal edge of mesepimeron and metapleuron not invaginated ( Figs. 20 View FIGURES 17–22 , 28 View FIGURES 23–28 , 37 View FIGURES 35–40 , 54 View FIGURES 53–58 ); 1 = lateral edge of pronotum and dorsal edge of mesepimeron and metapleuron invaginated, creating a longitudinal furrow ( Fig. 46 View FIGURES 41–46 , inv).
This invagination is present in Nosodipara and Pseudoceraphron , and is partially laterally concealed by the head (although visible ventrally) in all species except Nosodipara monteithorum .
51. Acropleuron: 0 = normal ( Figs. 20 View FIGURES 17–22 , 37 View FIGURES 35–40 , 46 View FIGURES 41–46 , 47 View FIGURES 47–52 ); 1 = large, convex, broadly expanded along entire dorsal length of mesopleuron ( Figs. 9, 10 View FIGURES 5–10 , 11 View FIGURES 11–16 , 28 View FIGURES 23–28 , 54, 55 View FIGURES 53–58 ).
Pyramidophoriella has a large, convex acropleuron which is not expanded anteriorly and is coded as ambiguous.
52. Posterior Mesopleural Sculpture: 0 = mesepipleuron posterior to femoral depression smooth; 1 = mesepipleuron posterior to femoral depression sculptured.
Although variable and difficult to code in many taxa, this character is included because it was found to reliably separates typical females of Dipara < Parurios > (smooth) from those of Dipara s. s. (rough). Mesolelaps and Pyramidophoriella have a strongly sculptured area surrounded by smooth areas, and are coded as ambiguous for this character.
53. Axillary Wing Sclerite: 0 = not visible; 1 = expanded, present as tegula-sized sclerite latero-posterior to mesowing bud ( Fig. 30 View FIGURES 29–34 , aws).
Myrmicolelaps paradoxus is unique in that the axillary wing sclerite is expanded and completely covers the mesowing bud.
54. Metatibial Spur Number: 0 = 2; 1 = 1.
It should be noted that the number of metatibial spurs in the Conophorisca + Myrmicolelaps clade is more variable than the taxa coded imply, as examination of undescribed species in the latter clade shows multiple reversals to 2 spurs. The number of metatibial spurs appears to have been reduced within many groups, including Cleonyminae ( Gibson 2003) , Toryminae ( Grissell 1995) , and Chalcididae ( Wijesekara 1997) .
55. Metatibial Spur Length: 0 = longest spur <1.5X as long as width of tibia at point of insertion ( Figs. 32, 33 View FIGURES 29–34 ); 1 = longest spur at least 2X as long as width of tibia at point of insertion (Fig. 98).
The longer metatibial spur in Pseudoceraphron < Dipareta > regieri is between 1.5–2X the width of the tibia, and is coded as ambiguous.
56. Metacoxal Striations: 0 = absent ( Figs. 23 View FIGURES 23–28 , 49 View FIGURES 47–52 ); 1 =present ( Figs. 21 View FIGURES 17–22 , 28 View FIGURES 23–28 , 32 View FIGURES 29–34 , 54 View FIGURES 53–58 ).
There are some genera in which different species have varying degrees of overall sculpture. In the case where many species in a single genus have strong, apparent striations and other have faint, less apparent strations, the taxon is coded 1 in this analysis.
57. Metacoxal Brush: 0 = absent ( Figs. 21 View FIGURES 17–22 , 28 View FIGURES 23–28 , 32 View FIGURES 29–34 , 46 View FIGURES 41–46 , 54 View FIGURES 53–58 ); 1 = present ( Fig. 39 View FIGURES 35–40 ).
A thick vertical brush of white setae is present on the posterior margin of the metacoxa in Neapterolelaps .
58. Metacoxal Shape: 0 = normal, posteriorly convex ( Figs. 21 View FIGURES 17–22 , 23, 28 View FIGURES 23–28 , 32 View FIGURES 29–34 , 39 View FIGURES 35–40 , 54 View FIGURES 53–58 ); 1 = laterally compressed, large concave surface facing posteriorly ( Fig. 49 View FIGURES 47–52 ).
59. Admarginal Setae (in winged forms): 0 = absent; 1 = present.
All winged Diparinae appear to possess longer than normal setae on the ventral surface of the forewing just posterior to the marginal vein. If the female is apterous, and the male is both known and winged, the taxon is coding according to the male. Darling (1991a) noted the presence of admarginal setae are also present in many eulophid genera.
60. Ratio of Marginal Vein to Stigmal Vein: 0 = marginal vein is less than 1.5X length of stigmal vein; 1 = marginal vein is greater than 3X length of stigmal vein.
61. Petiole Length (Female): 0 = broader than long to 1.2X as long as broad; 1 = at least 2X longer than broad.
62. Petiole Shape: 0 = cylindrical, 1 = strongly constricted antero-ventrally ( Figs. 11, 12 View FIGURES 11–16 , 34 View FIGURES 29–34 ); 2= midway along petiole length, curves downward at a sharp angle.
State 2 is autapomorphic for Conodipara .
63. Petiolar Setal Pairs (Female): 0 = petiole without setae ( Figs. 11, 12 View FIGURES 11–16 , 34 View FIGURES 29–34 ); 1 = petiole with 1 to 4 pairs of lateral or dorso-lateral setae; 2 = petiole with single pair of strong, dark bristles ( Fig. 53 View FIGURES 53–58 ).
State 2 is autapomorphic for Dipara < Pondia >. This state is added here instead of as a bristle character for two reasons. First, a unique bristle character for Dipara < Pondia > is autapomorphic and would be phylogenetically uninformative, and second, the bristle pair in Dipara < Pondia > is positionally homologous to the setal pairs in other taxa coded as state 1.
64. Petiolar Setal Tufts (Male): 0 = petiole without tufts of setae; 1 = petiole with lateral tufts of thick, white setae, at least in anterior half.
65. GT1 Constriction: 0 = GT1 rounded or straight lateral to petiolar insertion ( Fig. 12 View FIGURES 11–16 ); 1 = GT1 dorsoventrally compressed lateral to petiolar insertion, forming posteriorly extending carina ( Figs. 71, 72 View FIGURES 71–72 ).
66. GT1 Size: 0 = normal, covering less than a quarter of the distance from anterior margin of gaster to anterior margin of cercus; 1 = expanded, covering at least half of the distance from anterior margin of gaster to anterior margin of cercus ( Figs. 12 View FIGURES 11–16 , 34 View FIGURES 29–34 ).
Within Diparinae only Pyramidophoriella lacks an expanded GT1.
67. Setal Tufts on GT1: 0 = absent ( Figs. 11, 12 View FIGURES 11–16 , 19 View FIGURES 17–22 , 34 View FIGURES 29–34 ); 1 = present ( Figs. 37 View FIGURES 35–40 , 50, 51 View FIGURES 47–52 ).
When present, thicks tufts of long, white setae are present on the anterior surface of GT1 just lateral to the petiole.
68. Cercal Form: 0 = exerted, digitform; 1 = flat, plate-like.
Gibson (2003) coded most cleonymines as having plate-like cerci, although the hypothetically primitive genera Bouckekius and Chalcidiscelis have digitform cerci. Digitiform cerci have also been recorded in Archaeopelma (Eupelmidae) ( Gibson 1989), female Torymidae , Agaonidae s. l., and Chromeurytominae (Pteromalidae) ( Grissell 1995).
69. Cercal Setae: 0 = short; 1 = long ( Fig. 13 View FIGURES 11–16 , 17 View FIGURES 17–22 , 24 View FIGURES 23–28 , 34 View FIGURES 29–34 , 35 View FIGURES 35–40 , 41 View FIGURES 41–46 ).
70. Cercal Brush: 0 = absent ( Fig. 24 View FIGURES 23–28 ); 1 = present ( Figs. 13 View FIGURES 11–16 , 17 View FIGURES 17–22 , 34 View FIGURES 29–34 , 35 View FIGURES 35–40 , 41 View FIGURES 41–46 ).
The cercal brush is a dense patch of setae abutting the anterior rim of the cercus (see Methods section).
71. Sexual Dimorphism: 0 = absent; 1 = present.
Minor differences in metasomal and antennal shape are not coded as dimorphism. Conodipara , Conophorisca , and Myrmicolelaps were all coded as absent for this character. However, rather than possessing generalized females and males as Liepara and the outgroup taxa do, these African genera have males which resemble the specialized wingless females typical of diparines.
72. Flagellar Segments (Male): 0 = flagellar segments cylindrical and short, at most 1.5X as long as wide; 1 = flagellar segments cylindrical and long, at least 1.5X as long as wide; 2 = flagellar segments pedunculate and long, at least 1.5X as long as wide.
This character is coded only for sexually dimorphic genera with known males, so as not to add weight to the sexual dimorphism character itself.
73. Flagellar Setae (Male): 0 = male antennae with short setae; 1 = male antennae with long, erect setae; 2 = male antennae with long, apically appressed setae.
As with the previous character, this character is coded only for sexually dimorphic genera with known males.
74. Number of Anelli (Male): 0 = 1 anellus; 1 = 3 anelli.
This character is included in addition to sexual dimorphism because Spalangiopelta is sexually dimorphic and the male has 3 anelli, whereas in sexually dimorphic diparines, the male has only a single anellus regardless of the female. Characters #74–76 may appear out of order in a morphological sense. This is because they were added to the matrix after the initial matrix was completed to further elucidate the relationship between Ceinae and Diparinae .
75. Claval Peg-Like Sensilla: 0 = absent; 1 = present.
This character was identified by Darling (1991a) as a potential synapomorphy for Ceinae + Diparinae , although the genus Bohpa ( Darling 1991b) does not possess them.
76. Torular Position: 0 = antennal toruli separated from oral fossa by a distance of less than 1 torulus diameter; 1 = antennal toruli separated from oral fossa by a distance of greater than 2 torulus diameters ( Figs. 8 View FIGURES 5–10 , 16 View FIGURES 11–16 , 27 View FIGURES 23–28 , 36 View FIGURES 35–40 , 44 View FIGURES 41–46 ).
Phylogenetic methodology
Parsimony analyses were performed using PAUP* version 4 ( Swofford 2002). All characters were treated as unordered. Heuristic searches were conducted using 100 random addition sequences, and multiple states were treated as variable (ambiguous data were distinguished from polymorphic data). Characters were then reweighted using the rescaled consistency index, and a new heuristic search was performed. This process was repeated to determine if topology would continue to change with successive reweighting steps. Although there was minor variation in the number of steps following each reweighting/search iteration, tree topology never varied from the topology determined by the initial reweighting/search step. In all analyses the combined notauli character (#31) was used rather than the separate notaular characters (#32–34) to avoid overweighting of notauli. Two permutations of the data set were tested. First, Bohpa (Ceinae) was both included and excluded in analyses. Second, a data set was tested in which all bristle positional characters were removed (#22–30). The reasoning for these permutations is discussed in the previous section. Character state evolution was traced using MacClade, version 3 ( Maddison and Maddison 1992), and all indices were calculated using PAUP* version 4 ( Swofford 2002).
Phylogenetic results
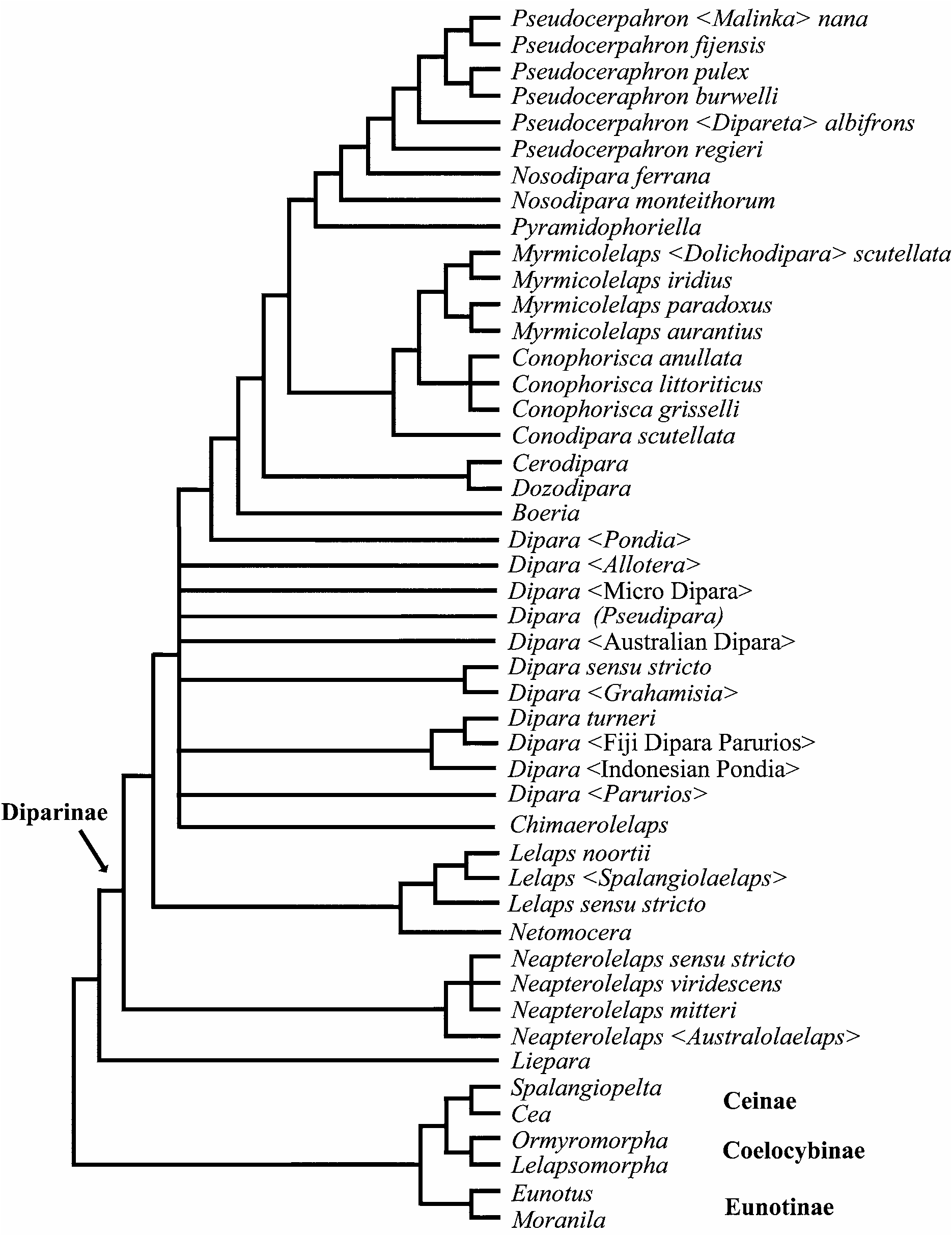
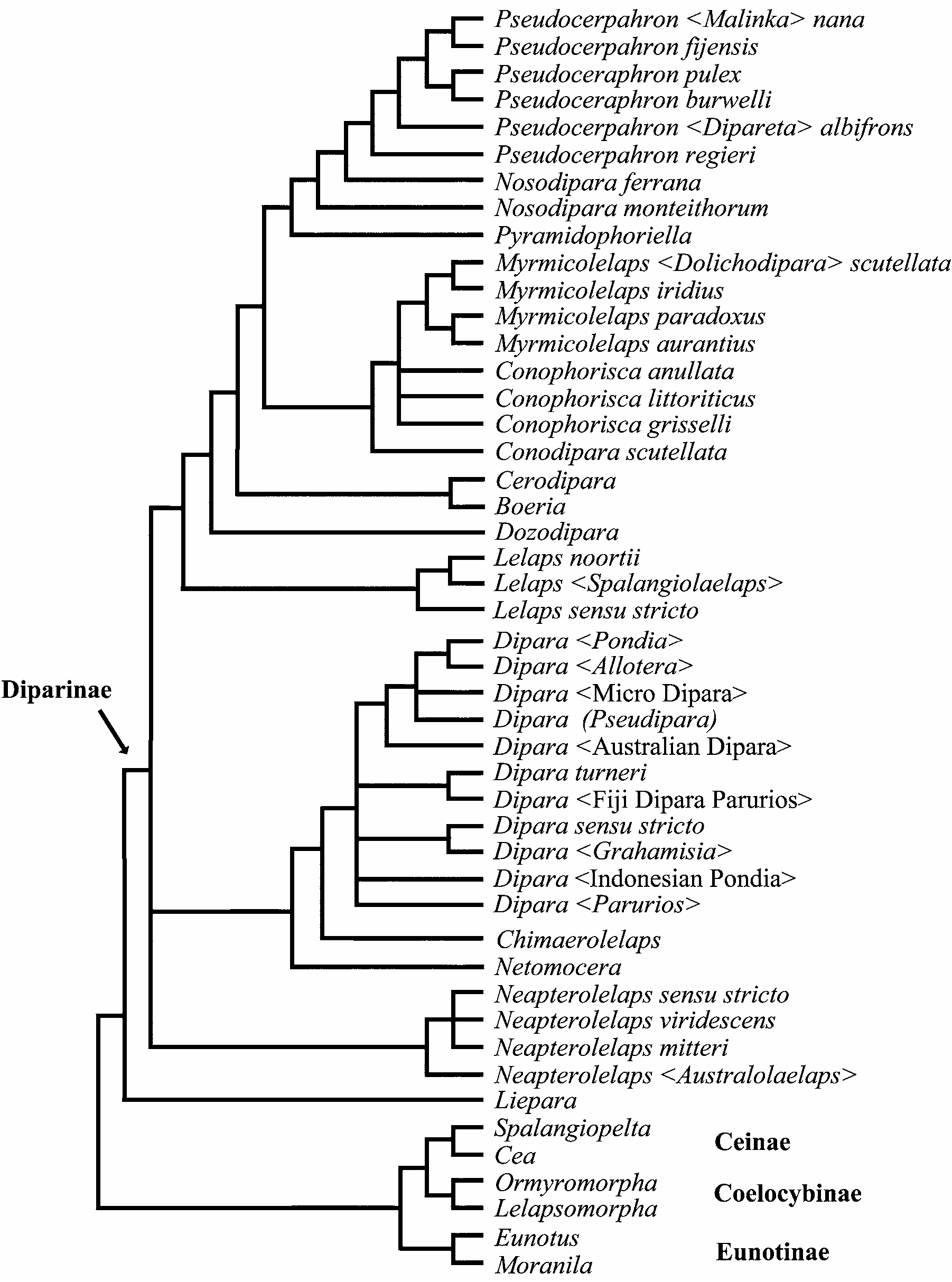
The parsimony analysis including Bohpa and bristle positional characters resulted in 125 trees (290 steps, not shown), and after the characters were reweighted, the heuristic search resulted in 45 trees (strict consensus shown in Figure 1 View FIGURE 1 ). The parsimony analysis including Bohpa but excluding bristles resulted in 2501 trees (243 steps, not shown). After the characters were reweighted, the heuristic search resulted in 511 trees (strict consensus shown in Figure 2 View FIGURE 2 ). The parsimony analysis excluding Bohpa but including bristle positional charac- ters resulted in 973 trees (280 steps, not shown), and after the characters were reweighted, the heuristic search resulted in 63 trees (strict consensus shown in Figure 3 View FIGURE 3 ). The parsimony analysis excluding Bohpa and bristle positional characters resulted in 127 trees (234 steps, not shown). After the characters were reweighted, the heuristic search resulted in 170 trees (strict consensus shown in Figure 4 View FIGURE 4 ).
Diparinae excluding Liepara Bouček , Eunotinae , Ceinae , and Coelocybinae was recovered as monophyletic in all analyses. In no analyses was Liepara recovered within Diparinae . Liepara appeared as sister-group to Diparinae when Bohpa was excluded. Ceinae was recovered as monophyletic in analyses excluding Bohpa , but paraphyletic as (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae in analyses including the taxon. Bohpa was recovered as sister-group to Diparinae in analyses in which it was included. In three of the analyses Neapterolelaps represented the basal divergence within Diparinae , and in the fourth analysis it formed a basal polytomy with (( Dipara + Chimaerolelaps ) + Netomocera ) and the more derived diparines. In the analysis including Bohpa and bristle positional characters, Lelaps was recovered as paraphyletic. In the remainder of the analyses it was monophyletic. Dipara was recovered as a paraphyletic grade in the analysis excluding Bohpa and including bristle positional characters, and in the remainder of the analyses was recovered as monophyletic. In all analyses Boeria , Cerodipara , and Dozodipara formed a paraphyletic grade at the base of the remaining diparines, although their positioning with respect to each other varied across analyses.
Data Set Permutations
Bristle characters did not appear to uniformly impact tree construction between analyses both including and excluding Bohpa . Character state reconstructions for the bristle positional characters were examined using MacClade ( Maddison and Maddison 1992), and different characters supported different clades. It is therefore unlikely that bristle positional characters present a single strong signal which is overwhelming other signal in the analyses with one major exeception. When bristle positional characters were included, there was a transformation requiring a large number of steps in the paraphyletic grade of Boeria and Cerodipara . Additionally, in the analysis including both Bohpa and bristle characters, Lelaps was recovered as paraphyletic at the base of this grade, most likely due to the absence of bristles in L. noortii . In analyses where bristles were excluded, Cerodipara and Boeria were sister-taxa and Lelaps was recovered as monophyletic. It therefore appears that bristles positively impact tree contruction in some areas and negatively in others.
The inclusion or exclusion of Bohpa had a dramatic impact on tree construction. When Bohpa was included it was sister-taxon to Diparinae , and this potential relationship is discussed below in the Sister-Group Relationships section. Additionally, the inclusion of Bohpa heavily influenced the internal phylogeny of Diparinae , most likely affecting the reweighting of characters important to the phylogeny of the derived diparines, because it had very little impact on which character states were ancestral for Diparinae . As Bohpa is flightless and appears highly modified for that role, it shares many characters with the more derived wingless diparines (e.g., a reduced metanotum), likely due to convergent evolution (in no analysis was Bohpa sistergroup to the more derived diparines). This caused many characters which appear to be of high phylogenetic utility in the other analyses to be strongly down-weighted in the reweighting step, as many characters became more homoplastic with Bohpa ’s inclusion (e.g., the inclusion of Bohpa in the analysis drops the RCI of the metanotum character (#41) from 0.63 to 0.44).
Evaluation of traditional characters
Although most diparines are easily recognized by chalcidologists, there has been no morphological evidence given to support their monophyly. They have traditionally been defined by various subsets of seven characters, although the phylogenetic value of these characters has not previously been examined. Additionally, these characters have only been used diagnostically in regional revisions, so their utility on a world level has also been unknown. The distribution of these characters throughout Diparinae and Pteromalidae is discussed below, and the phylogeny is used to evaluate their diagnostic utility.
1) Strong, dark bristles on the vertex and dorsal surface of the mesosoma (character #21). Although bristles are considered a diagnostic character for Diparinae , they are absent in many genera (e.g., Neapterolelaps , Myrmicolelaps ) and their presence and thickness is variable in others (e.g., Dipara ). Additionally, similar mesosomal bristles are present in many other pteromalids, including some eunotines, coelocybines, and species of Spalangiopelta (Ceinae) . Only the configuration of the vertex bristles appears to be unique among the diparines. In Diparinae , the vertex bristles are arranged along the occipital margin, ocellar triangle, and along the dorso-frontal margins of the eyes ( Fig. 18 View FIGURES 17–22 ). In other pteromalids with vertex bristles, the bristles are often uniformly distributed across the entire vertex (e.g., Liepara , Lelapsomorpha (Coelocybinae) , Spalangiopelta ( Ceinae )). Based on the phylogeny, bristle presence/absence appear to be extremely homoplastic (RCI = 0.06–0.07) within Diparinae and more broadly Pteromalidae , and provides little phylogenetic or diagnostic utility.
2) Transverse striations on the posterior margin of the metacoxa (character #56). Striations are present on most diparines, although they are absent in Pseudoceraphron and Nosodipara , and their strength can vary dramatically within genera. The presence of this character outside Diparinae is difficult to determine,
as many pteromalids have some sculpture on the posterior surface of the metacoxa, which is often difficult to judge whether or not it is transversely striate. This character has limited phylogenetic and diagnostic utility because of its sporadic loss within Diparinae (RCI = 0.14–0.19) and coding difficulties outside of the diparines. However, this character does support a close relationship between Diparinae + Liepara , with the character either evolving at the base of ( Diparinae + Liepara ) + Eunotinae and being subsequently lost in Moranila , or being independently derived in both Diparinae + Liepara and Eunotus .
3) High insertion of the metacoxa. Many diparines have their metacoxal insertion significantly higher than their mesocoxal insertion. However, this character shows continuous variation throughout Diparinae , and both coxae are inserted on the same level in some taxa (e.g., Dipara < Pseudipara >), making metacoxal insertion difficult to use as a diagnostic feature. This character was not coded for the phylogenetic analysis, because it could not be divided into discrete states.
4) Expanded first gastral tergite (GT1) covering at least a third of the metasoma (character #66). This character is present in all diparines except Pyramidophoriella , although it is also distributed throughout many pteromalid subfamilies (e.g., Eunotinae , some Miscogasterinae ). As with the metacoxal striations, this distribution limits the character’s diagnostic value. The ancestral condition within the Diparinae is an expanded GT1 (RCI = 0.20–0.38). An expanded GT1 appears to be sympleisiomorphic for Diparinae if Liepara is the sister-group, as it appears in the phylogeny to be synapomorphic for ( Diparinae + Liepara ) + Eunotinae . However, when Bohpa is sister-group to Diparinae , an expanded GT1 is synapomorphic for Diparinae . The expansion of GT1 may prove to have value in higher phylogenetic studies of Pteromalidae .
5) Antennae with one anellus, seven additional funicular segments, and three club segments (characters #15, 16, 74). This feature is broken up into three characters in the phylogenetic analysis: number of anelli in the female (#15), number of anelli in the male (#74), and number of flagellar segments in the female (#16). These characters have little value for diagnosing diparines, as many diparine females have more than one anellus (e.g., Pseudoceraphron ), and many other pteromalid groups have similar antennal formulas (e.g., some Cleonyminae ). The number of anelli in diparines appears to correlate with body size, as the most diminutive genera (e.g., Pseudoceraphron ) have the most anelli, and in Dipara an increase in anellar number appears to correlate with a decrease in body size. Although these characters have limited diagnostic value, they do provide useful phylogenetic information and all have high consistency indices (#15: RCI=0.55–0.72; #74: RCI=0.33–1.00; #16: RCI=1.00). Presence of a single anellus is pleisiomorphic for the Diparinae when Liepara is sister-taxon and synapomorphic when Bohpa is sister-taxon. An increase in anellar number is synapomorphic for the Nosodipara + Pseudoceraphron clade, and a second increase is synapomorphic for Pseudoceraphron . The number of anelli in both males and females support a sister-group relationship of Ceinae + Coelocybinae rather than Ceinae + Diparinae . A reduced number of flagellar segments is synapomorphic for Eunotinae .
6) Female often wingless. This character is extremely plastic within Diparinae at the genus, species, and intraspecies level, and was therefore not coded in the analysis. For example, many genera contain both macropterous and apterous species (e.g., Dipara and Lelaps ), and Bouček (1988) noted a single species of Australian Dipara which had females showing the full range of macroptery, brachyptery, and aptery. Other characters included in the analysis provide a more accurate and less variable measure of ‘potential’ wing size. For example, the posterior notal wing process (#39) is always present and fully expanded in genera which have both macropterous and apterous species, while it is often reduced or absent in genera which have only wingless species. The diagnostic utility of aptery is limited; many wingless female pteromalids are diparines, but many female diparines are not wingless.
7) Presence of long cercal setae (character #69). This character is present throughout Diparinae and Liepara , and is the only synapomorphy for the combined group (RCI=0.17–0.38). The reconstruction of long cercal setae is ambiguous when Bohpa is sister-taxon to Diparinae .
Monophyly of Diparinae
While monophyly of Diparinae is supported in all analyses, the synapomorphies which unite the clade differ whether Liepara or Bohpa is considered the sister-taxon. Only a single synapomorphy unites Diparinae in both analyses: the presence of a cercal brush. When Liepara is considered sister-taxon to Diparinae , diparine monophyly is supported by five additional synapomorphies: lack of a smooth, convex dorsellum, presence of sexual dimorphism, presence of an occipital carina, presence of admarginal setae, and a long marginal vein. When Bohpa is considered sister-taxon to Diparinae , diparine monophyly is supported by three additional synapomorphies: an expanded GT1, transverse striations of the posterior margin of the metacoxa, and a single anellus in the female. Diparinae is diagnosable by a combination of two of these features: lack of a smooth, convex dorsellum and presence of a cercal brush. Those characters which have not already been analyzed are discussed below.
1) Presence of a cercal brush (character #70). All diparines have a cercal brush (which is defined in the Methods section). The size and density of the brush varies throughout the Diparinae , but it is always present ( Figs. 13 View FIGURES 11–16 , 17 View FIGURES 17–22 , 34 View FIGURES 29–34 , 35 View FIGURES 35–40 , 41 View FIGURES 41–46 ). The cercal brush is synapomorphic for Diparinae in all analyses (RCI=1.00). In a broad survey of Pteromalidae , the cercal brush was only noted in two other taxa. First, Spalangia (Spalangiinae) possesses a similar brush. The spalangiines appear to be unrelated to the diparines, as they have little morphological similarity, and molecular evidence also suggests a lack of close relations (Desjardins et al. unpublished). The presence of this character in both taxa could represent convergent evolution, as both taxa inhabit similar environments. The only other genus in Spalangiinae , Playaspalangia , was not examined for this character. However, Playaspalangia is a rare genus known only from Mexico and Sri Lanka, which parasitizes Diptera under rotting algae ( Yoshimoto 1976), and may be a highly derived species of Spalangia . Additionally, a sparse cercal brush was found on the holotype of Spalangiopelta ferrierei , although it appeared absent on the two additional female specimens of the species. To assess the state of this character throughout Spalangiopelta , multiple specimens of 3 additional Spalangiopelta species were examined ( S. canadensis , S. ciliata , and S. felonia ). None of these species showed any evidence of a cercal brush, so the character is coded as absent in the genus, with the sparse brush in S. ferrierei being considered an aberrant condition.
2) Absence of a broad, convex dorsellum (character #42). All diparines lack a broad, convex, dorsellum ( Figs. 10 View FIGURES 5–10 , 19 View FIGURES 17–22 , 29 View FIGURES 29–34 , 38 View FIGURES 35–40 , 48, 50, 51 View FIGURES 47–52 , 55 View FIGURES 53–58 ) (RCI=1.00). In some diparines the metanotum is absent entirely, so the theoretical presence of a dorsellum cannot be ruled out in those taxa. However, the most primitive diparines do possess a metanotum which is not reduced, and lack a dorsellum, so it is unlikely that the dorsellum is “hypothetically regained” in taxa lacking a metanotum. A reduced, smooth metanotum without a convex dorsellum has been noted in other pteromalids, particularly Bohpa and an undescribed, apterous cleonymine from Madagascar. Additionally, this state is present in the apterous encyrtid Tetracyclos boreios (redescribed in detail by Gibson and Yoshimoto 1981). This reduction may be an adaptation for increased mesosomal mobility in apterous chalcidoids, and therefore may have been independently derived in Bohpa . This feature is synapomorphic for Diparinae when Liepara is its sister-taxon and Diparinae + Bohpa when Bohpa is its sister-taxon. Moranilines ( Eunotinae ) also lack a broad, convex, dorsellum, and instead have a cup-shaped projection issuing from the medial portion of the metanotum. However, this configuration is different from the undifferentiated metanotum in diparines and is coded as a separate state.
3) Sexually dimorphic (character #71). Most diparines with known males are sexually dimorphic, although this trait was lost once within the group (RCI=0.28). When Liepara is considered sister-taxon to Diparinae , sexual dimorphism evolves twice independently, once in Diparinae and once in Spalangiopelta . Alternatively, in analyses including Bohpa , sexual dimorphism evolves at the base of the Ceinae and is subsequently lost in Cea or evolves twice independently in Diparinae + Bohpa and Spalangiopelta . Despite the phylogenetic value of sexual dimorphism, it provides little diagnostic utility. Most often only single specimens are available for identification purposes, and even when multiple specimens are present, it is difficult to
identify them as con-generic or -specific.
4) Presence of an occipital carina (character #6). The presence of an occipital carina appears to be synapomorphic for Diparinae when Liepara is its sister-group (RCI=0.09–0.13), although in analyses including Bohpa , it evolves at the base of the Ceinae and is subsequently lost in Cea or evolves twice independently in Diparinae + Bohpa and Spalangiopelta . Its low consistency index and sporadic distribution throughout Pteromalidae make the occipital carina’s phylogenetic utility difficult to judge. A selection of outgroups which have occipital carinae would likely have given different results. The presence of an occipital carina has no diagnostic value for Diparinae , as the trait is transformed multiple times within the group.
5) Admarginal setae (character #59) and 6) Long marginal vein (character #60). These character are discussed below in the Sister-Group Relationships: Bohpa Darling section.
Sister-group relationships: Liepara Bouček
In his revision of the Australasian Chalcidoidea , Bouček (1988) synonymized the existing tribes of Diparinae and erected a new tribe within the subfamily, Lieparini , for the newly described genus Liepara . Although Bouček made no specific statement that Liepara was sister-group to the remainder of the Diparinae , his tribal classification suggests this relationship. Bouček united Liepara with the Diparinae based on five characters: 1) antennal formula 11173, with a clearly three-segmented symmetrical clava, 2) GT1 distinctly enlarged, 3) hind coxa inserted fairly high, 4) hind coxa with distinct transverse striations, and 5) typical diparine-like pattern of bristles. As discussed above, many of these characters have limited phylogenetic value, due to their sporadic presence throughout Pteromalidae (1, 2, 5) or coding difficulties (3, 4), and none of these characters appear to be synapomorphic for Diparinae + Liepara or place Liepara within Diparinae (with the possible exception of 4, discussed below). The pattern of vertex bristles is particularly interesting, because the interpretation of Liepara ’s vertex bristles here differs from that of Bouček (1988). As already noted, the vertex bristles of diparines are arranged along the occipital margin, ocellar triangle, and along the dorso-frontal margins of the eyes. The vertex bristles on Liepara appear to uniformly cover the vertex, and more closely resemble the bristles of Lelapsomorpha and Spalangiopelta than those of Diparinae . While Lelapsomorpha and Spalangiopelta appear to have a much greater number of bristles than Liepara , they also have a much smaller scrobe and larger vertex, providing more positionally homologous space for the placement of bristles.
Liepara does not appear to belong to Diparinae and in particular lacks the highly consistent cercal brush and a medially undifferentiated metanotum. Liepara appears as the sister-group to the remainder of Diparinae in phylogenetic analyses excluding Bohpa , although the only synapomorphy uniting this clade is long cercal setae (#69). The presence of metacoxal striations (#56) also supports this relationship in one of two most parsimonious reconstructions (the other reconstruction is ambiguous). Additionally, this sister-group relationship is supported by biogeography. The most basal lineage of diparines, the Neapterolelapini, has an Australasian distribution, as does Liepara . Netomocera , another primitive diparine, is also present in this region. Ceinae , on the other hand, which will be discussed below as another potential sister-group to Diparinae , is one of the few pteromalid subfamilies absent from the Australasian region.
Even if Liepara is sister-group to Diparinae as presented here, maintaining Liepara within Diparinae would greatly weaken the definition of the latter group. Two qualitative diagnostic features would be eliminated and replaced with a single character which is much more difficult to diagnose, as the length of the cercal setae is more quantitative than qualitative. Additionally, the support for Liepara as sister-group to Diparinae is not strong, as Bohpa is recovered as sister-group to Diparinae when included in the analysis. The taxonomic effects of removing Liepara from Diparinae and Liepara ’s placement within Pteromalidae are further discussed in the genus’ treatment in the generic revision.
Sister-group relationships: Bohpa Darling
Ceinae is a cosmopolitan (although absent from the Australasian region) pteromalid subfamily including three genera: Bohpa , Cea , and Spalangiopelta . In the phylogenetic analysis excluding Bohpa , Ceinae was resolved as monophyletic based on five synapomorphies, only the first of which is not homoplastic: presence of claval peg-like sensilla, first funicular segment subequal in length to pedicel, presence of admarginal setae, marginal vein long, and toruli within 1 torulus diameter from the oral fossa. In these analyses, Ceinae was placed as sister-group to Coelocybinae based on three synapomorphies: female with three anelli, male with three anelli, and GT1 not expanded. The number of anelli may not be independent between non-sexually dimorphic taxa, although Spalangiopelta shows both sexual dimorphism and three male anelli, while all diparines, whether sexually dimorphic or not, have only one anellus in the male.
When Bohpa is included, however, Ceinae appears paraphyletic as (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae . The entire clade is supported by two synapomorphies: the presence of admarginal setae and a long marginal vein. The reconstruction of sexual dimorphism is ambiguous in part because the male of Bohpa is unknown; in one reconstruction it is gained independently in Spalangiopelta and Bopha + Diparinae , in the second it is gained at the base of the entire clade and subsequently lost in Cea . Regardless, both reconstructions support a close relationship between Diparinae and Ceinae . The clade of Bohpa + Diparinae is supported by the loss of the dorsellum, and Spalangiopelta + Cea is supported by claval peg-like sensilla. Darling (1991a) discusses six characters traditionally used to define Ceinae , and their relevance to a mono- and paraphyletic Ceinae are discussed here.
1) Complete notauli. This state appears pleisiomorphic for both a mono- and paraphyletic Ceinae , as all taxa included in the analysis except for the coelocybines and Pyramidophoriella have complete notauli.
2) Propodeal spiracles positioned halfway along antero-posterior axis of propodeum. Darling mentions the presence of this character in some Colotrechninae . Whether this character state is present in some diparines depends on whether or not the nucha is included in the propodeal measurement. Some diparines with a long nucha, such as Myrmicolelaps , have a fairly posteriorly placed spiracle. If the propodeum is measured without the nucha, the spiracle is positioned medially. However, if the propodeum is measured with the nucha, the propodeal spiracle is positioned anteriorly. Regardless, this state would be derived within the diparines, as all basal diparines have anteriorly positioned spiracles. Although not coded in the analysis, this would support a monophyletic Ceinae .
3) Toruli situated within 1 torulus diameter from oral fossa. This state is only present in the derived diparine genera Boeria and Cerodipara . Darling mentions the lower positioning of the toruli occurs in some neodiparines, eunotines, cleonymines ( Graham 1969), and colotrechnines ( Bouček 1988). Within this phylogenetic analysis, torular position supports a monophyletic Ceinae when Bohpa is excluded, and is gained at the base of (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae and subsequently lost in Diparinae when Bohpa is included.
4) Antennal formula 11353. Darling (1991a) notes the presence of three anelli in some Pteromalinae , Miscogasterinae , and Pireninae . The presence of three anelli in both sexes is also found in Coelocybinae and when Bohpa is excluded is synapomorphic for Coelocybinae + Ceinae . However, when Bohpa is included, this feature is independently derived in Coelocybinae and (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae , and subsequently lost in Diparinae .
5) Marginal vein long. Darling does not discuss the distribution of this character throughout Pteromalidae , but this feature is synapomorphic for (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae when Bohpa is included. When Bohpa is excluded, a long marginal vein is independently derived in Diparinae and Ceinae .
6) Mandibles bidentate. As with the marginal vein, Darling does not discuss the distribution of this character. This character was not treated in the phylogenetic analysis due to the difficulty of coding many diparine genera known from only a few specimens whose mandibles could not be observed.
Prior to the description of Bohpa, Darling (1991a) hypothesized that ceines were monophyletic based on two putative synapomorphies, the first of which is a reduced number of papilliform sensilla. He hypothesized that Diparinae + Ceinae is defined by papilliform sensilla, and further that Ceinae is defined by a reduced number of those sensilla (alternatively, in an equally parsimonious explanation, Diparinae could be defined by an increased number of papilliform sensilla). As defined by Darling (1991a), papilliform sensilla are socketed, lobate setae that appear in pairs on the apical margin of the funicular segments in ceines. These structures were previously unknown in the Chalcidoidea . Some eulophids have more simple, unsocketed, randomly placed structures termed “multiporous pegs”, but Darling stated that the two are likely not homologous. Darling sampled representatives from 15 pteromalid subfamilies, and papilliform sensilla were found only on Lelaps , a member of Diparinae . Although such a complex character showed potential as a strong synapomorphy uniting ceines and diparines, Darling only investigated a single member of Diparinae . Unfortunately, this character could not be utilized in the phylogenetic analysis, as specimens of many diparine genera were unavailable for slide mounting or scanning electron microscopy. However, a variety of diparines were examined for this feature.
The antennae of males of four available diparine genera ( Lelaps , Netomocera , Dipara , and Neapterolelaps ) were slide mounted in Hoyer’s (results not shown). The slide of Lelaps was prepared as a positive control, in order the establish the position and visibility of the sensilla. Papilliform sensilla were found in all four genera (including the primitive Neapterolelaps ), suggesting that the presence of sensilla is the ancestral state for Diparinae . Antennal SEMs of the female of Pseudoceraphron (Diparinae) reveal a lack of the papilliform sensilla. This may be an artifact of the placement of papilliform sensilla, which in females are most often located beneath the dorsally extended apex of the multiporous plate sensilla. The female of Pseudoceraphron has a largely reduced funicle (7–8 segments are anelliform). Anelliform segments by definition lack multiporous plate sensilla, and therefore the majority of funicular segments in Pseudoceraphron lack a positionally homologous location for the placement of papilliform sensilla. As the male of Pseudoceraphron is not known for certain, it could not be examined for these sensilla. However, regardless of the distribution of papilliform sensilla across Diparinae , close investigation of antennal SEMs in Gibson’s revision of Eupelminae (1995) have revealed papilliform sensilla on a large number of eupelmids (e.g. Zaischnopsis , Fig. 354; Reikosiella , Figs. 367–368). Although this character may still prove useful in the phylogenetics of pteromalid subfamilies, its distribution must be more carefully examined.
Darling termed his second ceine synapomorphy “claval peg-like sensilla.” However, later that year he published the description of Bohpa (1991b) , in which he stated that Bohpa has neither the papilliform nor claval peg-like sensilla. Phylogenetic analyses in which Bohpa was included would suggest papilliform sensilla were lost in Bohpa , and claval peg-like sensilla are synapomorphic for Spalangiopelta + Cea . The exceptionally small size of Bohpa may have resulted in the reduction or loss of many features, including antennal ones. It is difficult to speculate on character evolution within Ceinae , however, as in a 3 taxon clade a gain and a loss are equally as parsimonious as two independent gains of a character state. Additionally, outgroups in this analysis were chosen to reflect possible diparine relationships rather than ceine ones.
In addition to the presence of papilliform sensilla, Darling (1991a) proposed the presence of admarginal setae on the inner leading margin of the forewing membrane as a synapomorphy for Ceinae + Diparinae . The presence of these setae does appear synapomorphic for (( Spalangiopelta + Cea ) + Bohpa ) + Diparinae in this phylogenetic analysis. Darling suggested that these setae might hold the hindwing tight against the body during movement in confined spaces, and during the course of this study, many diparine specimens were observed in which the setae appear to hold the hind wing in place while they are folded against the body. However, it should be noted that since both diparines and ceines search for hosts in leaf litter, this character could be a result of convergent evolution.
Inclusion of additional characters could help elucidate the relationship between Ceinae and Diparinae . The addition of a propodeal spiracle position would lend support to Ceine monophyly, while a papilliform sensilla character would lower support for a Bohpa + Diparinae relationship. However, knowledge of the male of Bohpa could potentially provide the strong support for a Bohpa + Diparinae relationship. If Bohpa is sexually dimorphic, this would support a close relationship between Ceinae and Diparinae , as the genus is likely sexually dimorphic. Also, if the male of Bohpa is winged and has admarginal setae and a long marginal vein, this could be the decisive evidence indicating Bohpa as the sister-taxon to Diparinae rather than Liepara . For now, however, the sister-group to Diparinae will have to remain in question.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.